1712
Логіка мислення. Зареєструватися 4. Підземна активність
Ми приїжджаємо до точки, де ми повинні переходити від розвідки основ нейронауки та теорії нейромереж до нового, що містить запропоновану модель. Для тих, хто тільки почав читання серії, я рекомендую починати з першої частини.
Повернемося до опису роботи реальних нейронів. Вивіски з деяких нейронів через їх осей переходять до вводів інших нейронів. У хімічній шпинделі медіатор звільняється, яка в залежності від типу синхронного, має або активуючу або гальмуючу дію на отримання нейрона. Чутливість шпинделя, яка може змінюватися, визначає внесок цього синапсу до загального збудження. Якщо загальний вплив перевищує певний поріг, то мембрана деполірована і нейрона генерує шип. Спірайт є одним імпульсом, тривалість якого і амплітуда не залежать від того, яка симпатична активність генерувала його.
Найлегша модель, натхненна імпульсною активністю нейрона, є моделлю порогу. У той же час, виходячи з того, що шип може бути порівнюється з бінарним сигналом, вважається, що вводи і вихід додачі беруть значення тільки 0 і 1. Якщо вводи такого формального нейрона дають імпульсний візерунок, який повторюється від такти до такти, то нейрон, в залежності від налаштування його ваги, необхідно або дати зворотний сигнал або мовчати. Це досить логічно – постійне вхідне зображення відповідає постійним результатом на виході.
Якщо ви намагаєтеся привезти модель поріг до реальності, то перше, що вам потрібно зробити, припустимо, що шаблон вхідної активності може бути строго синхронізований. Тобто сигнали в різних вводах можуть бути закодовані імпульсами, що мають кожну частоту. При цьому припущення не можна просто використовувати миттєву картину стану вводів. Вам потрібно буде вибрати проміжок часу і використовувати картину активності, накопичену за цей інтервал. На різних частотах вхідних сигналів за деякими інтервалами, вхідні імпульси формуватимуть достатні для активації нейрона, а на інших дають розряди, залишаючи нейрон неактивний. Таким чином, відповідь нейрона буде набута власної частоти, яка буде залежати від частоти вхідних сигналів і чутливості відповідних сипсів.
Така причина призводить до моделі нейрона як лінійного підсилювача, в якому рівень сигналів при входіх нейрона і його відповідь описуються не двома рівнями, але по реальних кількостях, які відповідають частотам походів. Перехід на лінійну дошку значно спрощує моделювання та частково забуває про початкову хімічну природу нервової активності.
Але все має початок. Для отримання інформації про введення мозку потрібні нейрони, які взаємодіють з зовнішнім світом. Чутливість цієї взаємодії дуже відрізняється від синоптичної чутливості. У сипсах кількість яких в одному нейроні можна вимірювати в десятках тисяч, медіатор звільняється від бульбашок з постійною потужністю. Мінімальний рівень викиду – це частина близько 7000 молекул. Сенсорні нейрони працюють з різними обсягами. Таким чином, візуальні палички активуються з буквально двох квантів світла, нейрон-рецептори золочної системи здатні виявити лише кілька молекул запаху речовини. Таку високу схильність досягається за допомогою механізмів посилення внутрішнього сигналу.
Взаємодія нейрона з навколишнім середовищем відбувається через молекули білків – рецептори, які дозволяють зовнішній хімічний вплив змінити стан всередині клітини. речовина, яка взаємодіє з певним типом рецептора називається лігандом. Для сиптичних рецепторів, ліганди є дуже нейротрансмітери, які дозволяють нейронну взаємодію.
Сусідні рецептори можуть підключитися до створення дімерів (рис. нижче), які в свою чергу поєднуються для формування рецептивних кластерів.
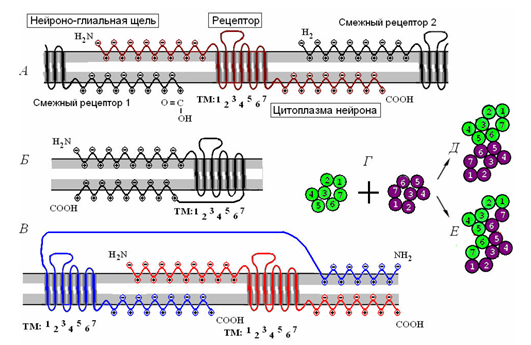
кластеризація рецептора. Це єдиний рецептор і його взаємодія з навколишніми рецепторами. B є мономерною молекулою рецептів. B - рецептивний димер. D - поєднання двох мономерів в контакт (D) і комбінації (E) (Радченко, 2007)
Рецептори діляться на іонтропні та метаботропні. Іонотропні рецептори створюють іонні канали, які переміщують заряджені іони по мембрані, змінюючи потенціал мембрани. Коли ліганд взаємодіє з іотропним рецептором, останні зміни провідності іонного каналу, відкриття або закриття його. Ці рецептори розташовані в сиптичних клацках. Їх спільна робота визначає, чи не існує адгезії, викликаних додаванням зовнішніх сигналів.
Трохи відрізняється природою в метаботропних рецепторів. Вони розташовуються поза спинами і не створюють іонних каналів. При взаємодії з лігандом вони запускають внутрішньоклітинні посередники, які посилюють початкові сигнали сотні тисяч разів, що в кінцевому рахунку призводить до активації нейрона. Це означає, що навіть невелика частка речовини, яка є лігандом для метаботропного рецептора може викликати відповідну нейрон для спіке. Цей механізм використовується за допомогою сенсорних нейронів, дозволяє їм реагувати на незначні зовнішні впливи. Але крім сенсорного сприйняття, метаботропні рецептори відповідають за значний відсоток діяльності мозку в цілому.
Характерний подразник для нейрона - це візерунок вхідних сигналів, що збігається з візерунком чутливості його приколів. Чим ближче вхідний сигнал полягає в характерному подразнику, тим вище частота походів, що створюються нейроном. Ця реакція називається індукованої діяльністю. У всіх нейронів, які якось реагують на поточний образ. Але індуковані активні рахунки тільки невелика частка загальної активності мозку. Основна активність падає на так звану фонову активність (рис. нижче).

Нейронська відповідь на стимули та фон (спонтанна) активність
Підземна активність складається з одноразових походів, що відбуваються з часу. Такі прикмети називають спонтанними, оскільки вони відбуваються незалежно від наявності або відсутності індукованої діяльності. Якщо ви захищаєте мозок від зовнішньої інформації, спонтанна активність в сенсорних зонах не ослабить, але навпаки, збільшиться тільки. Для індивідуального нейрона, його спонтанна активність виглядає як серія випадкових походів. Але для нейронів однієї кортильної зони ця активність додає до загального ритму, який разом створює значні електричні коливання. Це ці флуктуації, які можна зафіксувати шляхом застосування електродів до склепу, що фактично називається електроенцефалографії.
Для різних зон кори і різних станів людини характерні їх частоти і рівні таких ритмів. Найпотужніший ритм з окремої зони кори простежується на області шкіри безпосередньо над цією зоною. Тому для запису ритмів мозку використовують набір електродів рівномірно розподілений на поверхню голови. Результат виглядає як набір графіків, кожен письмовий з власних електродів (рис. нижче).
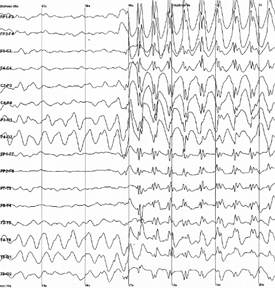
Приклад електроенцефалограми. різке збільшення амплітуди відповідає на початок епілептичного нападу
Якщо індукована активність є наслідком роботи іонтропних рецепторів, то спонтанна активність є результатом активації метаботропних рецепторів. Коли імпульси регулярної або спонтанної активності, що пропагують вздовж осі, досягають нейрона одержувача, вони викликають випуск посередника в сиптичну клітку. Цей медіатор визначає внесок синапсу до процесу провокованої нейронної активності. Потім відбувається його перезапуск, а синпс відновлюється до первісного стану. Але частина посередника виводиться за межі снаптики і поширюється через простір, утворений тілами нейронів і тілами навколишніх гліальних клітин. Це явище називається Spilover від англійського спілу - переповнення, переповнення. Поблизу сипси утворюють те, що називається сиптичних пасток. У цих пастках, інгібіторах та активуючих медіаторах створюють інтерференційний шаблон. Це означає, що кожне поєднання імпульсів викликає чіткість медіатора в певному унікальному місці мембрани. Радченко А.Н. показав, що метаботропні рецептивні кластери, розташовані в місцях таких нуклеї, здатні змінити свої властивості і згодом реагувати на повторення того ж зовнішнього імпульсу (Радченко, 2007).
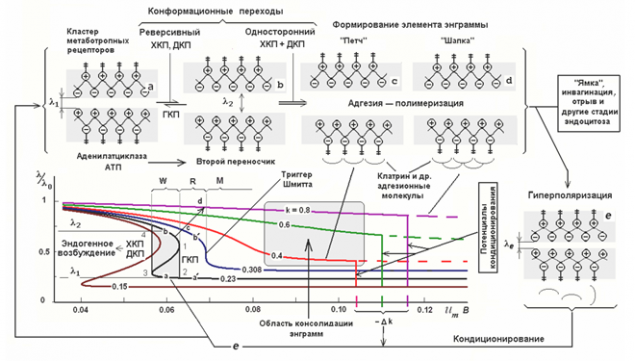 р.
р.Різні стани метаботропного рецептивного кластера (Радченко, 2007)
Радченко охарактеризував кілька станів, характерних для метаботропних рецептивних кластерів та у порівнянні з цими станами з різними стадями запам’ятовування. Уміння рецепторів впливати на стан клітинки залежить від положення їх сплітних кінців. Чим більша відстань між ними, тим сильніше зовнішні кінці протріть на поверхні, а внутрішні проникають в клітинне середовище. Відповідно, більша відстань відповідає більшій чутливості рецептора. У міру зменшення відстані торці мийки рецепторів в мембрану, а рецептори втрачають свою здатність впливати на стан нейрона. Дистанцію можна контролювати мембранним потенціалом нейрона. Деполяризація відштовхує кінці мембранних перегородок і в клітинку, відповідно, і гіперполяризація виводить їх назад в мембрану.
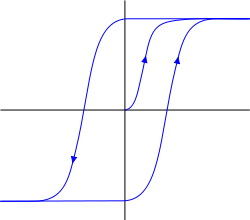
Спочатку метаботропні рецептивні кластери мають особливості гістерезу. Гістерез має на увазі, що поведінка системи визначається її предісторією (граф нижче). Після видалення сили, але залишається в цьому положенні. Щоб видалити її з цієї позиції, зараз необхідно застосувати зворотну силу, яка, після проходження певного порогу, переведена система на інший стабільний стан, що має однакову властивість «прилипання».
 Р
Ргістерез
Гістерез дає рецептори тригерні властивості. Реакція до чистоти медіатора, рецептори можуть «помакувати» як в стані, що відштовхується з мембрани, так і в стані занурення в ній. Перший стан говорить про те, що нейрон стає чутливим до певної картини оточуючої активності і може реагувати на її повторення власним спієм, другий, навпаки, робить його імунітетом до нього. Але це приклеювання є короткостроковим в природі, оскільки деякі зміни мембранного потенціалу можуть скидати кластер до його первісного стану.
Радченко в порівнянні з цією поведінкою рецептивних кластерів з короткостроковою пам'яттю. Що визначає нашу думку в будь-який момент, з одного боку, картина активності нейронів, а з іншого боку, фіксація слідів цієї діяльності, що дозволяє нам швидко відновити останні фотографії. Невагомість цих слідів забезпечує швидкість пам'яті. Ми можемо швидко повернутися до думки, що був другим або хвилиною тому, але ми втратимо цю простоту, коли години або дні проходять.
Під впливом медіаторів і сильного зсуву в мембранному потенціалі, рецептори можуть втратити свої властивості гістерезу і переходити в стабільний режим, який постійно фіксує придбаний стан кластеру, який можна порівняти з довгостроковою пам'яттю. У той же час, в залежності від положення фіксації, відштовхується стан або посухи, кластер або реагує на певну картину активності, або навпаки – набуває стабільну нечутливість до неї. Ці стабільні стани можуть зберігатися протягом тривалого часу, можна сказати назавжди, хоча в певних умовах вони можуть перезаписуватися.
Далі в нашій моделі ми припустимо, що мозкові нейрони мають три форми зберігання інформації. Однією формою є зміна чутливості сонок, що визначає характерний образ нейрона. Друга форма – короткочасна фіксація герметизації на метаботропних рецептивних кластерах картини навколишньої активності. А третя тривала фіксація, завдяки переходу рецептурних кластерів до стабільного стану, частин короткочасних фіксованих зображень.
Чутливість сипсів нейрона описує єдиний характерний образ (за винятком нейронів з декількома секціями сипсів). Додатковосинаптична поверхня нейрона, яка містить сотні тисяч метаботропних рецептивних кластерів, може зберігати порівняну кількість спогадів, що вказують на які події цей характерний образ проявляється самостійно.
Зверніть увагу, що імпульсна активність нейронів складається з двох компонентів. Один компонент - це патерн індукованої активності, який розповідає вам, які нейрони навчилися їх характерним стимулям. Друга складова – картина умовно спонтанної активності, яка існує відносно незалежно від першого, і що створює загальний фон мозку. Цей фон менш акцентовано, ніж індукована активність, але більш широкий, оскільки він розподіляється по всій кори.
Я не боїться сказати, що фонова активність є однією з найбільших міфів мозку. У наступній частині ми спробуємо вирішити цю таємницю.
Продовження
Література використана
Якщо щось занадто короткий, незнімний або непристойний, будь ласка, відписатися в коментарях. Моє око вже так розмитне, що мені важко покласти себе на місці когось, хто вперше сприймає цей матеріал. У той же час, якщо не складно, вказати рівень вашого загального знайомства з темою.
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/214525/
















Логіка мислення. Зареєструватися 3. Перцептрон, забруднювальні мережі
Логіка мислення. Зареєструватися 5. Латунні хвилі