1738
The logic of thinking. Part 4. Background activity
We have reached a point when retelling the basics of neuroscience and neural network theory we have to go to the new that contains the proposed model. For those who just started reading this series, I recommend starting with the first part of .
Let us return to the description of the work of real neurons. Signals from one neuron through their axons applied to the inputs of other neurons. In chemical mediator release occurs synapses, which, depending on the type of synapse is either the activating or inhibiting effect on the signal receiving neuron. Sensitivity of the synapse, which can vary, determined by the contribution of this synapse in the general excitement. If the net effect exceeds a certain threshold, then there is membrane depolarization and neuron generates a spike. Spike - a single pulse, the duration and amplitude of which does not depend on whether it has generated synaptic activity.
The simplest model, inspired by the impulse activity of neurons - is a model of a threshold adder. At the same time, based on the fact that the spike can be compared with a binary signal, it is believed that the inputs and output of the adder only take the values 0 and 1. If the inputs of the formal neuron impulse to submit a picture of a repetitive cycle in tact, the neuron as a function of adjust its weights must either every clock cycle to issue a response signal, or to remain silent. This is logical - a constant input pattern corresponds to a constant output results.
If you try to bring the model to reality threshold adder, the first thing that you have to do - is to assume that the input pattern of activity may not be strictly synchronous. That is, the signals on different inputs can be coded pulses, each having its own frequency. With this assumption, we can not just use an instant picture of the inputs. Need to pick a show time interval and use accumulated during this interval painting activity. At different frequencies of the input signals at certain intervals of the input pulses will form accuracy sufficient for activation of the neuron, and on the other to give discharge, leaving a neuron inactive. Thus, the response of the neuron acquires its own frequency, which will depend on the frequency of the input signals and the corresponding synapse sensitivity.
These considerations lead us to the neuron model as a linear combiner in which the signal level at the inputs of the neuron and its response are not described two levels and real values that correspond to the repetition frequency of spikes. The transition to a linear combiner allows greatly simplify modeling and partly to forget about the original chemical nature of neural activity.
But everything has its beginning. That the information entered into the brain, neurons are necessary, interacting with the outside world. The sensitivity of this interaction is very different from the synaptic sensitivity. At synapses, the number of which one neuron can be measured in tens of thousands, the mediator released from vesicles having a constant capacity. The minimum quantum of neurotransmitter release - is the portion of the order of 7000 molecules. Sensory neurons are working all other volumes. Thus, visual literally sticks are activated by two quanta of light receptor neurons of the olfactory system can detect only a few molecules of odorous substances. This high sensitivity is achieved by internal signal amplification.
Neuron interaction with the environment is due to protein molecules - receptors that allow external chemical resistance to change state within the cell. A substance which interacts with a particular receptor type, referred to their ligand. For synaptic receptor ligands - these are the neurotransmitters that enable communication of neurons.
Nearby receptors can be connected, creating dimers (figure below), which, in turn, merge to form a receptive clusters.
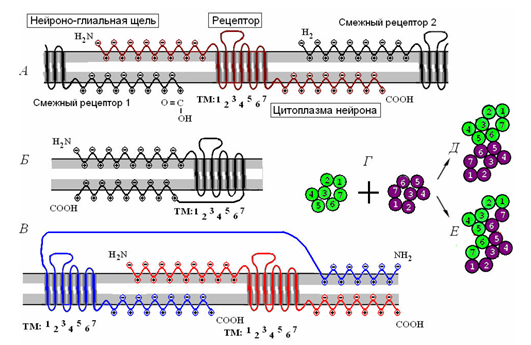
Clustering receptors. A - single receptor and its interaction with the receptor. B - monomer molecule receptive. B - receptive dimer. T - association of two monomers in contact (D) and the combination (E) dimers. (Radchenko, 2007) i>
Receptors are divided into ionotropic and metabotropic. The ionotropic receptors create ion channels which move the charged ions across the membrane, changing the membrane potential. When the ligand interacts with ionotropic receptor, the last change ion channel conductivity, opening or closing it. It is these receptors are located in the synaptic clefts. Their joint work determines to be or not to be a spike caused by the addition of external signals.
A somewhat different nature from the metabotropic receptors. They are located outside of synapses and create ion channels. When interacting with its ligand they start work intracellular mediators that provide amplification of the original signal in the hundreds of thousands of times, which ultimately leads to the activation of the neuron. This means that even a small proportion of a substance which is a ligand for metabotropic receptors, may cause a corresponding neuron spike. This mechanism uses sensory neurons, it allows them to respond to third-party effects insignificant. But in addition to sensory perception and metabotropic receptors are responsible for a significant proportion of brain activity at all.
Typical incentive for neuron - is this pattern of input signals, which coincides with the picture of the sensitivity of its synapses. The closer to the input characteristic of the stimulus, the higher frequency spikes generated neuron. This reaction is called induced activity. It occurs in all those neurons that once responded to the current image. But due to the activity is only a small part of the overall activity of the brain. But the main activity falls on the so-called background activity (see Figure below).

The response of the neuron to a stimulus and background (spontaneous) activity i>
Background activity is made up of emerging from time to time single spikes. Such adhesions are called spontaneous as they appear regardless of the presence or absence of induced activity. If you protect the brain from external information, spontaneous activity in the sensory areas will not weaken, but on the contrary, will only intensify. For a single neuron his spontaneous activity looks series of random spikes. But for one cortex neurons that activity consists in the general rhythm that creates significant total electrical oscillations. It is these fluctuations can be fixed by attaching electrodes to the scalp, which in fact is called electroencephalography.
For different areas of the cortex and the various states rights inherent in their frequency and levels of such rhythms. The strongest rhythm from a separate area of the cortex can be traced on the skin, which is located directly above this area. Therefore, to record brain rhythms using a set of electrodes, evenly distributed over the surface of the head. The result looks like a set of graphs, each of which is recorded on its electrode (see Figure below).
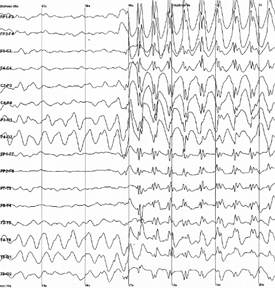
Example electroencephalogram. The sharp increase in the amplitude corresponds to the onset of an epileptic seizure i>
If the called activity - a consequence of the work of ionotropic receptors, the spontaneous activity - the result of activation of metabotropic receptors. When pulses are regular or spontaneous activity, spreading along the axon, the neuron reach the recipient, they cause the release of neurotransmitter into the synaptic cleft. This mediator determines the contribution of the synapse in the process caused by the activity of the neuron. Then there is its reuptake, and synapse is restored to its original state. But part of the mediator is expelled outside the synaptic cleft and spreads over the space formed by the bodies of neurons and their bodies surrounding glial cells. This phenomenon is called spillover from the English spillover - overflow, overflow. Located close to each other synapses form so-called synaptic traps. These traps brake and activating mediators create an interference pattern. This means that each combination of pulses causes antinode mediator in a certain unique position of the membrane. AN Radchenko showed that metabotropic receptive clusters are located in places such anti-nodes are capable of changing their properties and subsequently respond to the repetition of the same environmental pulse pattern (Radchenko, 2007).
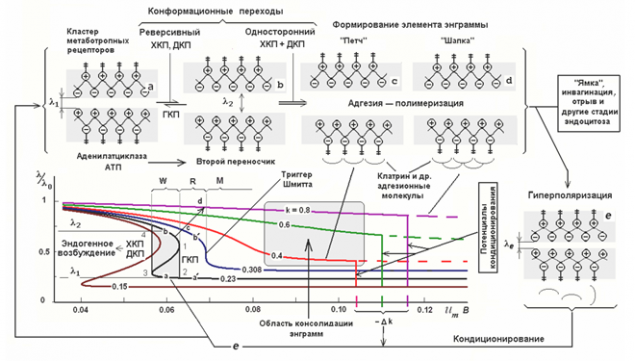
The various states of metabotropic receptive cluster (Radchenko, 2007) i>
Radchenko described several conditions that characterize metabotropic receptive clusters and compared these states with different stages of storage (picture above). The ability of receptors influence the state of the cells depends on the position of the split ends. The greater the distance between them, the more the outer end of the projecting surface, and penetrate into the internal environment of the cell. Accordingly, the greater the distance corresponds to the high sensitivity of the receptor. When reducing the distance the ends immersed in the membrane receptors, and receptors lose their ability to influence the state of the neuron. The distance may be controlled by the membrane potential of the neuron. Depolarization of the end portions pushes out from the membrane and into the cell, respectively, and oppositely hyperpolarization draws them into the membrane.
Originally metabotropic receptive clusters have hysteresis characteristics. Hysteresis implies that the behavior of the system is determined by its prehistory (chart below). Turning to the definition of sustainable saturated state, the system comes back after removal efforts, and remains in this position. To remove it from this position must now attach the opposite force that after passing a certain threshold bring the system to another stable state, has the same property "sticking».
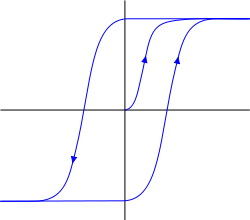
Hysteresis i>
Hysteresis gives receptors trigger properties. In response to an antinode neurotransmitter receptors can be "sticky" in the ejected state of the membrane, and in a state immersed in it. The first state indicates that the neuron becomes sensitive to certain environmental picture activity and can respond to its own repetition spike and the second, on the contrary, it makes it insensitive. But it is short-term stuck, as a definite change in membrane potential can reset the cluster to its original state.
Radchenko contrasted this behavior receptive clusters with short-term (short-term) memory. That is to say that at any given time determines our thinking - is, on the one hand, the pattern of neuronal activity, and on the other hand, such a fixation traces of this activity, which allows you to quickly recover recent pictures. The fragility of these tracks provides RAM. We can quickly return to the idea, which was a second or a minute ago, but lose this lightness, when they pass hours or days.
Under the influence of mediators and a strong shift in the membrane potential receptors may lose hysteresis properties and transition to a stable regime that permanently fixes the acquired state of the cluster, which can be compared with long-term memory. Thus, depending on the position of locking recessed push condition or cluster or responding to the activity of a certain picture or vice versa - becomes stable insensitive thereto. These stable states can be stored for a long time, we can say forever, although under certain conditions, they can be overwritten.
Further, in our model, we assume that the brain's neurons have three forms of information storage. One form - a change in the sensitivity of synapses, which determines the typical image of a neuron. The second form - a short-term trigger lock on metabotropic receptive clusters painting environmental activity. And the third - a long-term fix, due to the transition receptive clusters in a steady state, part briefly recorded pictures.
The sensitivity of the neuron synapse describes only his characteristic way (if we ignore the neurons with several sections of synapses). In this extrasynaptic surface of the neuron that contains hundreds of thousands of metabotropic receptive clusters can store a comparable number of memories, indicating in any event shows the characteristic image.
Please note that the impulse activity of neurons consists of two components. One component - this is the picture evoked activity, which suggests that some neurons have learned their characteristic incentives. The second component - a picture conditionally spontaneous activity, that there is a relatively independent of the first, and that creates a general background of the brain. This background is less accentuated than that caused by the activity, but more colossal for as distributed over the entire space of the cortex.
I am not afraid to say what the background activity - one of the biggest mysteries of the brain. In the next section we will try to give the answer to this mystery.
Continued
References
If there is too short, it is not clear or vague stated, please unsubscribe in the comments. I already have the eye so zamylen that I find it hard to put yourself in the place of the one who first sees this stuff. At the same time, if not difficult to select your level of familiarity with the general theme.
Previous parts:
Part 1. Neuron
Part 2. Factors
Part 3: Perceptron, convolutional network
Alex Redozubov (2014)
Source: habrahabr.ru/post/214525/
















The logic of thinking. Part 3: Perceptron, convolutional network
The logic of thinking. Part 5. Brain Waves