1708
思维的逻辑。第4部分后台活动
我们已经达到了一个点时,神经科学和神经网络理论,我们有基本的复述去到新的包含了模型。对于那些谁刚开始读的系列,我建议你开始的 中的第一部分。
让我们回到现实神经元的描述。从通过它们的轴突的神经元中的一个的信号被施加到其它神经元的输入。在化学突触释放介体,这取决于突触的类型或者是活化或在接收到的神经元的信号抑制效果。灵敏度的突触,其可以变化,由突触的一般兴奋的贡献来确定的。如果总曝光超过某一阈值时,膜去极化产生的神经元产生一个尖峰。穗 - 是一个单脉冲,持续时间和幅度,其中不依赖于哪个突触活动产生它
。
最简单的模型,灵感来自神经元的冲动行为 - 是加法器的门限的典范。在这种情况下,假设该尖峰可与二进制信号进行比较,可以相信,该加法器的输入端和输出端仅取数值0和1。如果一个正式的神经元输出的脉冲模式的输入,从循环重复,以时钟周期,神经元,这取决于调整其权重各周期必须或者发出一个响应信号,或者是沉默的。这是符合逻辑的 - 直流输入模式对应于一个恒定的输出结果
。
如果您尝试将模型到加法器的门限的现实,你必须做的第一件事 - 就是假定该活动的输入模式可能不是严格同步。也就是说,在不同的输入信号可以被编码的脉冲,每一个都具有其自身的频率。与此假设不再能够简单地使用的输入的状态的瞬时图像。需要选择一个显示时间槽,这个区间的绘画活动中使用累积。在以一定间隔的输入脉冲的输入信号的不同频率将形成精度足以使神经元的激活,而在另一给放电,而使不活动的神经元。因此,神经元的响应获取它自己的频率,这将依赖于输入信号和相应的突触的灵敏度的频率。
这种考虑导致我们对一个神经元的模型作为一个线性组合器,其中所述信号中的神经元和其响应的输入端的电平不是由两个层次,并且真实数量相对应的尖峰的重复频率进行说明。到线性组合器的过渡可以大大简化建模和部分忘记的神经活动的原始化学性质。
但是任何事物都有它的开始。该信息输入到大脑,需要神经元,与外界交互。这种相互作用的灵敏度是从突触敏感度非常不同。在突触,其中有一个单独的神经元的数目可以在数万被测量,神经递质从与永久容量小泡释放。神经递质释放的最小量子 - 7000分子的顺序的一部分。感觉神经元工作的所有其他卷。因此,视觉枝硬是用嗅觉系统的光受体神经元的两个量子激活可以检测的恶臭物质只有几个分子。这个高灵敏度由内部信号放大来实现的。
与环境的神经元的相互作用是由于蛋白质分子 - 受体使细胞内的外部化学品暴露变化状态。的物质,其与受体的一种特定类型的交互时,称他们的配体。对于突触受体的配体 - 这些是神经递质,使神经元的相互作用
。
毗邻的受体可以被连接,产生二聚体(如下所示),这反过来又合并,以形成一个接受集群。
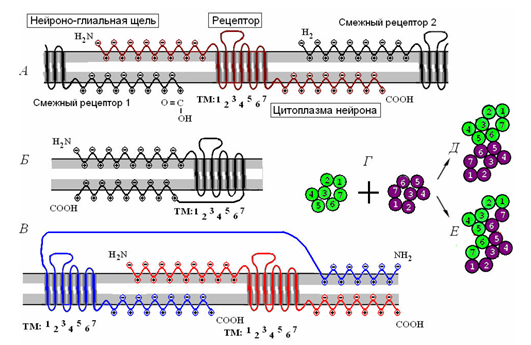
受体的聚集。 A - 单一受体和其与周围的受体相互作用。 B - 接受单体分子。在 - 接受二聚体。的T - 两种单体的接触(D)工会和组合(E)的二聚体。 (拉琴科,2007年) I>
受体分为离子型和代谢型。的离子型受体创造离子通道,在带电离子穿过细胞膜,改变膜电位。当配位体与离子型受体相互作用,后者改变离子通道的开口的导电率或关闭它。这些受体位于突触间隙。他们的联合工作决定生存还是毁灭尖峰造成的增加外部信号。
一个稍微不同性质的代谢型受体。它们存在于突触的外面,并创建离子通道。当与其配体相互作用后,便开始工作的胞内介体提供原始信号的放大在几十万次,这最终导致神经元的激活。这意味着一种物质,它是一种配体,代谢型受体的甚至一小部分时,可引起相应神经元的峰值。该机制使用的感觉神经元,它允许他们到一个可怜的第三方效应作出反应。但除了感觉和代谢型受体是负责一个显著的份额大脑活动的。
典型刺激神经元 - 是输入信号的一个图象,它与它的突触敏感度的图形一致。越接近输入信号的特性的刺激,频率较高的尖峰所产生的神经元。该反应被称为诱导活性。它发生在所有这些神经元以某种方式反应,以当前图像。但由于大脑的全部活动的一小部分的活性。但主要的活动落在了所谓的背景活动(见下图)。

神经元的刺激和背景(自发)活性 i>的
响应
背景活动是由不时出现以时间单次尖峰。这样的粘连称为自发因为它们出现无论存在或不存在诱导活性的。如果保护大脑免受外部的信息,自发活动在感觉区没有削弱,而是只会加剧。对于单个神经元的自发活动看起来像一系列的随机尖峰。但一个皮层神经元,本次活动是由在创建显著的总电振荡的总体节奏。正是这些波动可以是固定的通过将电极贴在头皮上,这实际上被称为脑电图。
对于不同的皮层区域和人性的不同状态,以自己的频率和节奏等的水平。最重的节奏在皮肤上,这是位于这个区域的正上方的区域中观察到的皮质的分开的区域。所以,用于记录使用一组电极脑节律,均匀地分布在头部的表面上。结果看起来像一组曲线图,其中每一个被记录在您的电极(见下图)。
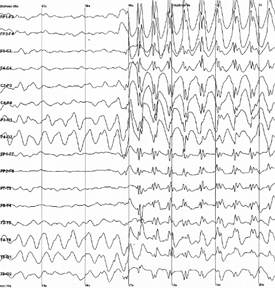
例脑电图。在振幅急剧增加对应的癫痫发作 i>的
发作
如果被调用的活动 - 它是一个后果的促离子型受体,自发活动 - 活化代谢型受体的结果。当脉冲是规则的或自发活动,沿轴突扩展,神经元到达接收方,它们会导致神经递质释放到突触间隙。这个介体确定到突触诱导神经元活动的过程中的贡献。再有就是它的再摄取,和突触被恢复到其原始状态。但中介的一部分被排出突触间隙外利差过的机构和周围的神经胶质细胞神经元体形成的空间。这种现象被称为溢出英语溢出 - 溢出,溢出。位于彼此接近,以形成所谓的突触,突触陷阱。在这些陷阱的制动和激活介质创建的干涉图案。这意味着,脉冲的每个组合产生波腹介体膜中的一些独特的位置。 AN拉琴科表明,位于地代谢容易接受簇如腹是能够改变它们的性质,并随后在相同的环境脉冲图案(拉琴科,2007)的重复响应的。
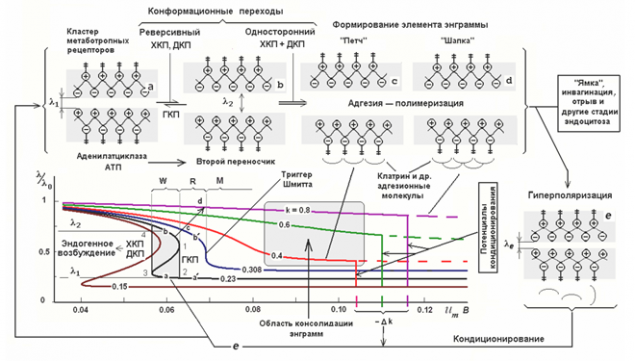
在集群的不同代谢接受状态(拉琴科,2007年) I>
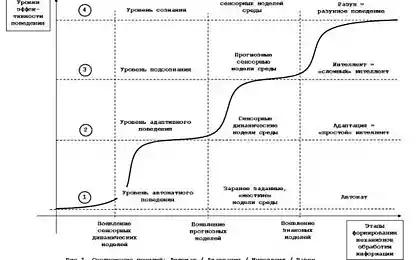
拉琴科描述的几种状态代谢接受集群的特点,并比较与存储的各个阶段(上图)这些条件。受体的能力影响的小区的状态依赖于分割端部的位置。它们之间的距离越大,越外端突出的表面上并渗透到细胞内部环境。相应地,更大的距离对应于该受体的高灵敏度。同的距离减小的端部浸渍在膜受体,并且受体的丧失来影响神经元的状态的能力。的距离可以由神经元的膜电位来控制。消偏推该膜的端部向外和向内分别的单元格,以及超极化计数器绘制成的膜。
最初代谢容易接受簇显示出滞后特性。滞后意味着该系统的行为是由其史前(下图)来确定。谈到可持续饱和状态的定义,系统恢复拆除的努力,并保持在这个位置。从这个位置取出她现在必须作出相反的力量,通过一定的阈值后使系统进入另一个稳定状态,具有相同的属性“置顶»。
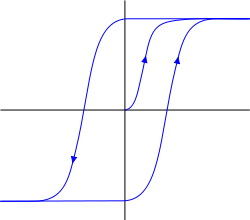
迟滞 I>
迟滞高度受体触发的属性。响应于波腹神经递质受体可能只有被排出的从膜和能够深入到它的“粘性”的状态。第一状态表示该神经元变得对某些环境图像活性敏感,并且可以响应它自己的重复尖峰和第二的,相反的,使得它不敏感。但它是短期的粘附,由于在膜电位的某些变化可以重置群集到其原始状态。
拉琴科对比与集群接受短期(短期)内存这种行为。这就是说,在任何给定的时间决定了我们的思想 - 是,一方面,神经元活动的模式,在另一方面,本次活动,它允许您快速恢复到最近的画作如此固定的痕迹。这些曲目的脆弱性提供了RAM。我们可以很快回到这是第二个或一分钟前的想法,但失去了易用性测试时,数小时或数天。
根据介质的影响和中膜电位的受体可能失去磁滞特性和过渡到稳定的制度以永久固定的群集,它可以与长期记忆进行比较所获取的状态的强烈变化。因此,根据锁定推压条件或凹陷,簇或响应于某一图片或反之亦然的活动性的位置 - 得到不敏感于此稳定。这些稳定状态可以存储很长,可以一直说,虽然在一定条件下它们可以被覆盖。
另外,在我们的模型中,我们假设大脑的神经元有三种形式的信息存储。一种形式 - 在突触的定义神经元的特征性图像的灵敏度的变化。第二种形式 - 这是对代谢接受集群环境绘画活动短期扳机锁。和所述第三 - 一个长期的固定,由于接受簇处于稳定状态过渡,部分简要地拍摄的图片
。
突触神经元的敏感性仅介绍了他特有的方式(除非你认为神经元突触的几个部分)。神经元,其中包含成千上万的代谢接受集群可以存储记忆相当的数量,说明在此情况下的这样突触外表面显示的特征图像。
需要注意的是神经元的冲动行为由两部分组成。一个部件 - 这是一个图象诱发活性,这表明神经元已经学会它们的特征的激励。第二组分 - 这是存在有条件自发性活动的图象是相对独立于第一,并创建大脑活动的一种常见的模式。这个图案被较少突出超过诱发活性,但更规模以及分布在皮层的整个空间。
不要犹豫地说,它是后台活动 - 大脑的最大谜团之一。在下一节中,我们将尝试给出一个线索,这个谜。
续
参考
如果某个地方过于简略,不清晰或难以理解的规定,退订请在评论中。我已经有了眼睛是如此模糊,很难把自己放在谁第一次看到这个东西的人的地方。同时,如果没有困难,说明你熟悉的题材一般水平。
前面的部分:
第1部分神经元
第2部分因素
第3部分:感知器,卷积网络
阿列克谢Redozubov (2014)
资料来源: habrahabr.ru/post/214525/