2279
Логіка мислення. Зареєструватися 5. Латунні хвилі

Отже, ми прийшли до опису одного з ключових принципів описаної моделі. Цей принцип не раніше був використаний в нейромережах або в описі як працює мозок. У зв'язку з цим я дуже рекомендую читати попередні частини. На мінімумі необхідно прочитати четверту частину, без якої описано нижче, буде повністю незрозумілим.
У попередній частині ми говорили про те, що активність нейронів ділиться на назву і фон. За допомогою електроенцефалограми спостерігаються види фонової активності. Зафіксовані сигнали мають складну форму і залежать від місця застосування електродів до голови, але, тим не менш, окремі гармонічні компоненти чітко простежуються в них.
Основні ритми назвали:
- альфа-ритм (від 8 до 13 Гц);
- ритм бета (від 15 до 35 Гц);
- ритм гамми (від 35 до 100 Гц);
- дельта ритм (від 0.5 до 4 Гц);
- Ата ритм (від 5 до 7 Гц);
- ритм sigma "spine" (від 13 до 14 Гц).
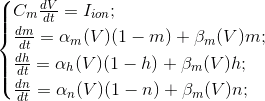
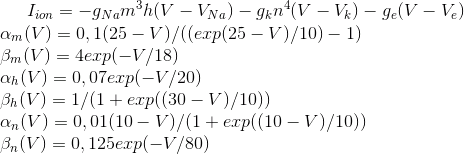
Ці рівняння визначають автоматичний процес, тобто вони призначають нейрон для створення імпульсів. Модель Hodgkin-Huxley важко моделювати. Таким чином, є багато спрощення, які зберігають основні генеруючі властивості. Найпопулярніші моделі Фітжуг (1961), Морріс С., Лекар Х. (1981), Гондмарш Ю. Л., Роже Р. М. (1984). Багато моделей, наприклад, Hindmarsh-Rose, дозволяють імітувати обидві дії пакету і ізольовані адгезії (рис нижче).
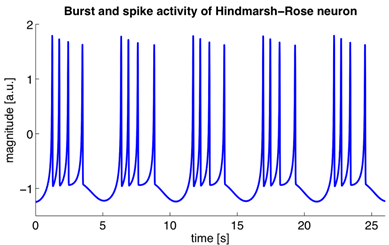
Пакетна активність та адгезії, які відбуваються при моделюванні нейрона
Поєднуючи нейрони, які генерують власні імпульси в дизайні, що нагадує структуру реальної кори, можна відтворювати різні ефекти, характерні для групової активності реальних нейронів. Наприклад, можна досягти глобальної синхронізації нейроактивності або викликати зовнішній вигляд хвиль. Найвідоміші моделі H.R. Wilson і J.D. Cowan (1972) і Kuromoto (1984).
Електроенцефалограмні записи лунок спільної активності нейронів, але зрозуміло, що ця активність має певну локацію часу. Методи оптичного спостереження за діяльністю кори дозволяють побачити його жити. У експериментальному тварині піддається зона кори і вводиться спеціальний барвник, який чутливий до змін електричного потенціалу. Під впливом загальної флуктуації мембранного потенціалу нейронів, такий барвник змінює спектральні властивості. І хоча ці зміни є надзвичайно малими, вони не можуть бути записані, наприклад, за допомогою діодного масиву, що діє як швидкісна відеокамера. Оптичні методи не дозволяють виглядати глибоко в корі і слідувати активності індивідуальних нейронів, але дозволяють сформувати загальну ідею перебігу хвильових процесів на її поверхні.
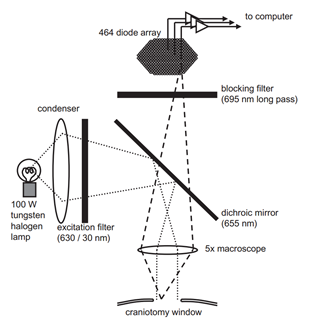 Р
РСхема оптичної фіксації кортичної активності (Михайл Т. Ліпперт, Кентаро Такагаки, Вейфенг Xu, Сяйоринг Хуан, Янь-Юнг Ву, 2007)
Виявилося, що ритми мозку відповідають хвилям, які відбуваються в точкових джерелах і пропагують через кору, як кола на воді. Однак передня хвиля на водних двержках строго по колу, а передня хвиля активність нейронів в мозку може поширюватися більш складним способом. На малюнку нижче показано візерунки пропагації хвилі на 5 міліметрової області кори щура.
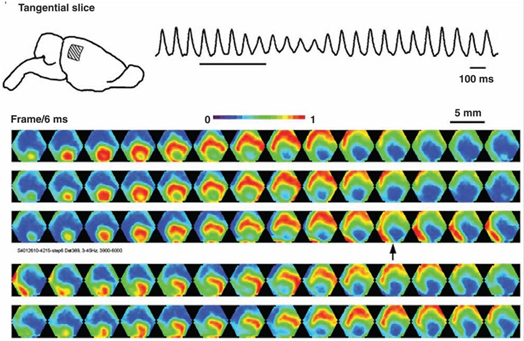
Картина поширення хвилі активності в області кори щура. Потенціал показаний градієнтом з синього до червоного. 14 кадрів в інтервалах 6 мілісекунди охоплюють один цикл пропагації хвилі (84 мілісекунди – 12 Гц) (Михайл Т. Ліпперт, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu, 2007)
Дуже цікаві та ілюстраційні відео хвильової активності даються в роботі (W.-F. Х.-Й. Гуан, К. Такагаки, Ю.-Ю. Ву, 2007. Виявляє, що хвилі можуть згубитися, досягаючи межі зони скоринки, можуть відображатися з іншої зони і створити протихвильову хвилю, може пропагувати в подвійних спіралі і створювати вихори.
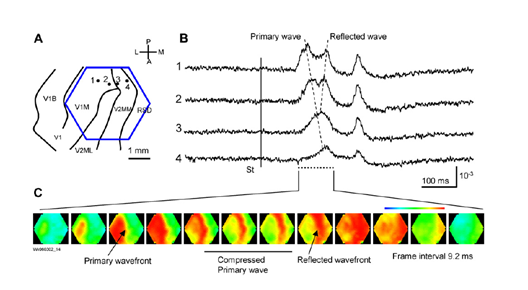
Пригнічення і відображення хвиль в первинній візуальній кори (W.-F. Xu, X.-Y. Гуан, К. Такагаки, Ю.-Ю. Ву, 2007)
Нижче відео з тієї ж роботи (W.-F. Xu, X.-Y. Huang, K. Takagaki, J.-Y. Wu, 2007):
Використовуючи моделі коливань нейронів, можна отримати аналогічні хвилі в комп'ютерних експериментах. Але сама по собі така пульсація не має сенсу. Важко припустити, що хвилі діяльності є механізмом передачі та обробки інформації. Але спроби пояснити характер цього механізму в традиційних моделях не виводяться відчутні результати. Здається логічно припустити, що інформація зашифрована частотою і фазовими характеристиками нейросигналів. Але це призводить до необхідності врахувати процеси інтерференції і підвищує більше питань, ніж уточнення.
Є гіпотеза про те, що хвилі «скану» скоринки, дані про читання з неї за її подальшу передачу (Питтс В., Маккулох В.С., 1947). Він виглядає досить здоровим, хоча б відносно альфа хвилі і первинної візуальної кори. Авторами цієї гіпотези Маккулок і Піттса підкреслюють, що сканування дозволяє пояснити одне важливе явище. Справа в тому, що інформація з первинної візуальної кори проводиться далі по волокнах білого матерії через пучки аксонів, явно недостатньо в обсязі для одночасної передачі всього стану цієї зони кори. Таким чином, вони укладаються, сканування дозволяє використовувати не тільки просторовий, але і часовий код, що забезпечує послідовну передачу необхідної інформації.
Ця проблема вузькості проекційних балок існує не тільки для первинної візуальної кори, але і для всіх інших ділянок мозку. Кількість волокон в проекційних шляхах набагато менша, ніж кількість нейронів, які утворюють просторий візерунок активності. З'єднання між зон чітко не здатні передавати весь просторово розподілений сигнал паралельно, що означає, що розуміння того, як інформація, що передається через них, стиснена і декомпресована. Витрата сканування, хоча це не відповідає на питання самого кодування, проте не дозволяє запитати правильні питання.
Наша модель в пояснюванні ритмів мозку ґрунтується на наведеному описі природи метаботропної активності. За допомогою ідеї, що не тільки сипси нейронів, але і метаботропних рецептивних кластерів беруть участь в процесі створення ритмів, дозволяє нам отримати результат, який якісно відрізняється від усіх класичних теорій. Але перед тим, що я хочу дати вам попередження. Я свідомо описую спрощуючі, ідеалізовані моделі, вирівняючи їх з певними процесами, які властиві реальному мозку, але не претендує на те, як працює мозок. Наше завдання полягає в тому, щоб показати основні принципи, розуміння, що еволюція є далеко вперед і їх істинна реалізація набагато більше хитрості. А аналогія може бути складена з розвитком обчислень. Сучасний комп'ютер досить складний, і якщо ми починаємо описати основні принципи класичних обчислювальних пристроїв, то виходить, що в чистому вигляді вони практично неможливо знайти в сучасних системах. Основне представлення – процесор перевіряє програму та дані з пам’яті, виконує дії, передбачені програмою на даних, та записує результати назад до пам’яті. Тепер додайте до цього використання різних рівнів кешу, багатопрочитаних, гіпертових, паралельних обчислень з використанням локальних, групових і спільних пам'яті, і таких як. І виходить, що важко знайти в реальному комп'ютері літрове дотримання простих правил. Насправді, все це необхідно враховувати, порівнюючи наступний опис з роботою реального мозку.
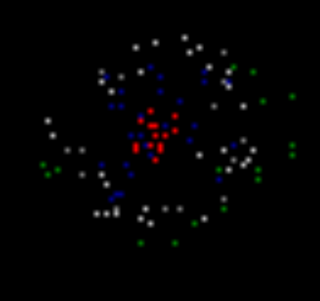
Отже, беремо модель кортичного регіону, де створюємо компактний візерунок індукованої активності. Повернемо питання про те, як сталося цей візерунок. Припустимо, що є елементи, на які є постійний імпульсний сигнал. На малюнку нижче нейрони, які утворюють цей візерунок, позначений червоним кольором. У реальній корі, це відповідає аксонам, що передає активність пачок, тобто видає ряд пайок з високою частотою. Ці осі можуть належати до нейронів в тій же області кори в стані індукованого збудження, або вони можуть бути проекційні волокна, що надходять з інших частин мозку.

Візерунок індукованої активності
Тепер ми отримуємо безкоштовні нейрони для створення рідкісних випадкових імпульсів. В той же час, ми накладаємо стан, який необхідний певний рівень навколишньої активності для випадкового проходу. Це означає, що випадкові адгезії можуть відбуватися лише поблизу вже активних нейронів. У нашому випадку вони з'являться в районі патерна індукованої активності. На малюнку нижче, випадкові шипи зображені в зелені.
Нормально, всі нервові дії, які не запускаються, називають спонтанною або фоновою діяльністю. Це не дуже успішним, оскільки він регулює сприйняття всіх цих дій як випадково. Далі ми покажемо, в загальному, фонова активність суворо заданий і не на всіх випадкових. І тільки невелика частина її дійсно повністю випадкові палички. Цей випадковий адгезії ми зараз створюємо навколо активного шаблону.
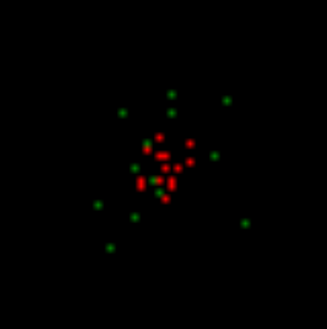
Перший крок моделювання. Спонтанна активність на тлі індукованої активності
Ми моделюємо метаботропні рецептивні кластери на наших формальних нейронах. Щоб це зробити, дозволяють нейронам запам'ятати, при необхідності картина активності найближчого середовища. І не обмежуйте себе однією картиною для одного нейрона, так як це буде, якщо ми запам'ятовували на сонці, але нехай кожен нейрон зберігає багато таких зображень.
Давайте зробимо це так, щоб присутність власної випадкової спиші та високої неоднорідності буде кожен раз, щоб запам'ятати картину навколишніх імпульсів (рис. нижче). Далі наші нейрони подарують один раз один з цих місцевих спогадів повторюється. Для того, щоб не заплутувати їх з випадкових спій, ми будемо називати ці походи хвилі.

Площа фіксації активності для одного з нейронів, які давали спонтанну косу (в центрі квадрата). У симуляторі використовуються квадратні приймальні поля для простоти, що, хоча це не відповідає полям відстеження реальних нейронів, не якісно впливає на результат.
В результаті в наступному кроці моделювання (фото нижче), ми отримуємо картину, в якій будуть однакові, як і раніше, нейрони з індукованої діяльністю (червоні), нейрони, які відповіли місцевим зображенням, що запам'ятовується в попередньому кроці (білий), а нейрони, які створювали спонтанний шип (зелений).
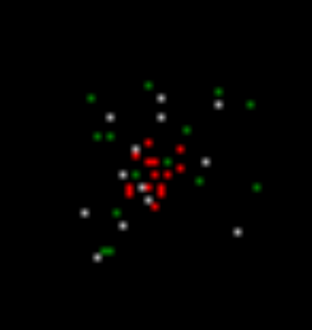
Другий крок моделювання. Червоний - індукована активність, білий - хвиля активність, зелений - спонтанна активність
Після повторення симуляційних кроків ми отримуємо активність, що поширюється через кору з певним унікальним випадковим чином створеним малюнком.
Тепер введемо стан нервової втоми. Давайте зробимо це так, щоб після декількох метаботропних (хвильових) шипок, нейрони втрачають здатність генерувати нові імпульси для часу, необхідний для відпочинку. Це призведе до того, що активність не буде пропагувати безперервною зоною, але шляхом пульсуючих хвиль, що працюють з джерела.
Щоб запобігти «зніманню», блокувати спонтанну активність на ділянках, де є велика кількість розслаблюючих нейронів. В результаті ми отримуємо кільце нейронів з хвилеподібною активністю, що працює від паттерна індукованої активності (рис. нижче).
Хвиля переднього поширення. Сині нейрони в стані релаксу.
Отримана хвиля, як вона рухається від центру, втратить щільність передньої і рано чи пізно буде згасати. На малюнку нижче показано перший цикл життя.
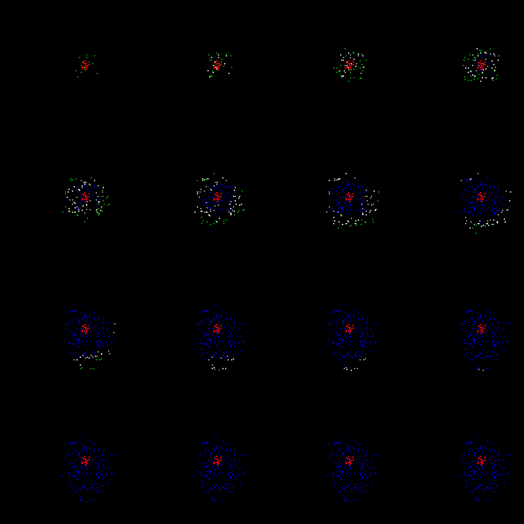
Перший цикл поширення хвилі ID
Після того, як пройшов період релаксації, стартує нова хвиля. Тепер буде підібрано нейрони, які вже пройшли навчання і розширюють трохи далі. З кожним новим циклом хвиля буде розширювати її діапазон до тих пір, поки вона не досягне країв зони. Щоб запобігти необмеженому збільшенню щільності хвиль, ми введемо інший обмеження: заборона спонтанної активності для нейронів, коли загальна активність навколо них перевищує певний поріг.
Через деякий час ми отримаємо кори, які навчаються пропагувати унікальний візерунок хвилі активності, що відповідає початковому даній схемі індукованої активності (рис. нижче).
1 999 р.
Пропагація хвиль за вже підготовленою кори (після 200 циклів навчання)
Нижче відео процесу навчання хвилі:
Зауважте, що коли кора вже навчається пропагувати хвилю з певного шаблону, зникають спонтанні адгезії. Таку «експериментальну» хвилю формується за рахунок хвилі, непередбачених походів. Випадкові оголошення з'являються тільки під час тренувань, коли фронт хвилі досягає непідготовлених або слабо підготовлених територій. На цій точці спонтанні імпульси створюють випадкове продовження хвильового малюнка, яке відразу пам'ятається на поверхні нейронів, залучених до цієї хвилі. Але, як тільки закінчилися тренування, хвиля від звичного шаблону починає поширюватися по вже створеному шаблону хвилі, кожен бар повторює точно так само, що вже не схожий на його шлях.
Кортикальне навчання не повинно бути поступовим. При моделюванні вибору параметрів можна досягти, що виникла хвиля є самодостатньою. Ось так, щоб він не згасився на перших стовпчиках, але відразу поширюється на весь простір (рис нижче).
1990Р. 2199Р.
Приклад невибагливості на першому циклі поширення хвилі навчання
Так як ширина хвилі фронту є постійним, його площа збільшується, так як вона рухається від джерела. Це відповідає тому, що більше і більше нейронів залучені до розподілу діяльності. Якщо ви простежуєте загальний потенціал, створений ними, ви отримаєте графіки, що ми бачимо на енциклограмах (рис. нижче).

Графік ритмічної активності єдиної моделі джерела
Зауважте, що ритми, що спостерігаються на таких «енцефалограмах», не є ритмом «поклоніння» кори. Що таке, що не синхронізуючий суглоб лопає активність, як очікується в більшості існуючих моделей, але змінюючи кількість нейронів, залучених до поширення диверенційних хвиль. Якщо зникнуть нові хвилі навіть перед фронтами попередніх хвиль, їх додавання додасть більш гладку картину. При певному підборі параметрів на моделі відтворюються ситуації, коли поширення хвиль практично не супроводжується загальною ритмічною активністю. Це може бути пов'язано з тим, що близько 10 відсотків людей на електроенцефалограмах хвильова активність не простежується.
Якщо ми зараз беремо новий візерунок індукованої активності, то скоринка створить хвилі, які пропагують з неї. Крім того, такий скорин навчається створювати хвилі для будь-якого стійкого малюнка індукованої активності. У зв'язку з тим, що цей самий нейрон може зберігати безліч локальних зображень, він може бути частиною багатьох різних хвиль, що відповідають різним шаблонам. Якщо ми бажаємо більшої унікальності хвиль, досить зменшити ймовірність його спонтанного шику, оскільки нейрон накопичує локальні зображення, що запам'ятовується.
Для пам'ятних локальних зображень важливо ввести механізм консолідації. Це, не зафіксуйте образ відразу протягом століть, але встановіть час, коли зображення потрібно повторити певну кількість разів. Так як підготовка хвилі та формування стабільних закономірностей індукованої активності є паралельними процесами, консолідація може витирати сліди невдалого навчання.
Але найголовніше у всіх це унікальність кожної з отриманих хвильових сюжетів. У будь-якому місці, через який проходить хвиля, він створює унікальний візерунок, характерний для нього. Це означає, що якщо є патерн провокованої активності на поверхні кори, яка зашифрує захід, після чого хвиля, викликана ним, буде поширювати інформацію про це по всій кортиці. У кожному конкретному місці ця хвиля дозволить створити свій унікальний візерунок, унікальний тільки для цього заходу. Будь-який інший захід дозволить створити в цьому місці інший візерунок, пов'язаний з його хвилями. Це означає, що в будь-якому місці ми можемо налаштовувати в «відновлення» хвилі від конкретної події і зафіксувати, коли буде відбуватися, коли хвиля прибуває з відповідним малюнком.
У нашій моделі виходить, що фундаментальні властивості кори:
- Уміння генерувати хвилі, які дивляться з візерунків індукованої активності
- вміння навчатися, проводити унікальний і стійкий до кожного малюнка хвиль;
- навчаються, здатність пропагувати хвилі активності, що виникають навколо стійких закономірностей індукованої діяльності.
Тепер процес навчання може бути грубо представлений наступним чином:
- статистично стабільні сигнали призводять до утворення нейронів детектора, які через сиптичну пластичність навчаються реагувати на певні візерунки;
- Детективні нейрони, що відносяться до одного шаблону, утворюють стабільний візерунок.
- Кожен стійкий візерунок, як це активний, вчить кору, щоб пропагувати унікальну хвилю, відповідну тільки до цього шаблону, яка може тлумачитися як ідентифікатор цього шаблону.
- Кожне місце земної кори, де відповідні хвилі пропагують, отримує можливість дізнатися, що на даний момент знаходиться загальна інформація.
Коли ми говоримо про структури мозку, які відповідають певним явищам, є очевидний дуалізму в нашій концепції. Ці структури не описуються виключно будь-якими нейронами або групами нейронів, але доповнюються ідентифікаційні хвилі, які дозволяють відповідну інформацію про поширення. Але, цікаво, в кожному конкретному місці, через яке хвиля проходить, вона поводиться таким же чином, як малюнок, який його створив. У моментально-часовому зрізі, це сам візерунок, який генерує продовження хвилі.
Не складно побачити, що це поширення ідентифікаційних хвиль повторює принцип Huygens-Fresnel, який описує поширення світлових хвиль. Суть його полягає в тому, що кожна точка фронту світлової хвилі може розглядатися як центр вторинного порушення. Кожен центр виробляє вторинні сферичні хвилі. Одержане світло поле є перешкодою цих хвиль. Конверт вторинних хвиль стає фронтом хвилі в найближчий час, і так далі.
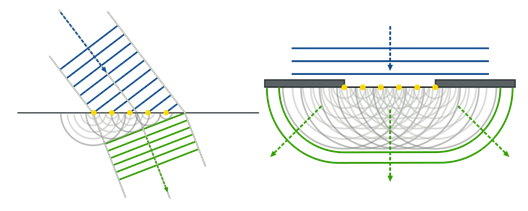
Рефракція (лівий) та втручання (праворуч) за даними Huygens-Fresnel (Nordmann)
Різниця в характері патерна індукованої активності і патерна, яка виникає під час пропагації хвилі може бути виявлена тільки шляхом аналізу причини активності нейронів, що надходять їх. Для індукованої діяльності це візерунок на сніпсах, для паттерна розмноження, реакція метаботропних рецепторів. Тобто відбиття мозку будь-яких явищ є як візерунок, так і хвиля, і в будь-який час, коли ми намагаємося виправити поширення цієї хвилі, ми побачимо візерунок, який відрізняється в шаблоні, але не сутність від того, що давало підніматися до хвилі.
Легко бачити, що дуалізм описується досить точно до дуалізму елементарних частинок. Кожна частинка - як частинка, так і хвиля. Залежно від того, які обставини ми розглянемо, частка може експонувати як хвиля, так і корпускулярні властивості. Сучасна фізика стверджує цей факт, але не пояснює його. Що ми описали про подвійність інформаційних образів, на щастя, дещо чіткіше, ніж дуалізм елементарних частинок. Питання, звичайно, чи можна продовжити ці аналогії і використовувати для розуміння фізичного малюнка світу. Це дуже серйозне питання, а пізніше ми будемо говорити про ДТП або неточність такого збігу.
Від дуалізму інформаційних зображень є важливі наслідки. Подібні частинки з різними хребтами, два візерунки з різних хвиль можуть бути в тому ж просторому регіоні без втручання один з одним. Крім того, кожна з них викликає продовження власної хвилі без введення взаємоперетворень. Якщо ми створюємо кілька візерунків і навчаємо кору пропагувати хвилі від кожного, то, активуючи ці візерунки разом, ми можемо спостерігати проходження хвиль через один одного, зберігаючи свою унікальність.
203481
Моделювання проходження хвиль з двох джерел
Це дуже відрізняється від безперервних хвильових процесів. Наприклад, в пропагації хвиль в воді або повітря всі молекули беруть участь в коливаннях. Зіткнення двох хвиль впливає на весь обсяг контакту, що призводить до перешкод. Поведінка кожної молекули визначається дією обох хвиль. Результатом є інтерференційний візерунок, який вже не так просто, як кожен візерунок хвилі індивідуально. У нашому випадку кожен з хвиль пропагує, впливає тільки на невелику частку всіх доступних нейронів. Коли дві довжини хвилі інформації міжсектифікують, нейрони вони діляться занадто мало, щоб заважати один одному. В результаті хвилі зберігають інформаційну картину, нерозриваючись від зустрічі з іншою хвилею.
Визначена елегантність описаної системи також полягає в тому, що нейрони можуть виконувати дві функції одночасно. Кожен нейрон може бути детектором, який налаштовується, змінюючи снаптичні ваги до певного образу. У той же час, у зв’язку з інформацією, записаною на його метаботропних кластерах, можна взяти участь у пропагації різних інформаційних хвиль.
Варто відчути різницю між властивостями нейрона як детектора, так і як учасник поширення хвильової активності. Здавалося б, і там він реагує на певну картину своєї діяльності. Насправді це два принципово різні властивості. Відносно гладкий тюнінг сиптичних ваг дозволяє нейрон вчитися ізолювати приховані фактори. Миттєва фіксація зображень за допомогою додаткових синоптичних кластерів забезпечує пам'ять про те, що зараз. Синапси сприймають великогабаритну картину з усього рецептивного поля нейрона. Метаботропні рецептивні кластери мають доступ до обрізного шаблону активності, обмежених сипперами, які утворюють їх сиптичними пастками. Комплект сиптичних ваг в звичайній нейроні є тільки одним, але він може зберігати десятки і сотні тисяч ідентифікаційних зображень. Синаптичне визнання генерує індуковану активність, тобто довгий пакет імпульсів. Кілька детекторних нейронів створюють візерунок індукованої активності. Метаботропне визнання дає єдиний спрей. Поєднання таких спійок створює ідентифікаційний фронт хвилі.
У моделюванні спостерігається, що діапазон пропагації ідентифікатора залежить від того, як активно проявляється малюнок, що проявляється. Застосування механізму консолідації, тобто, введення критеріїв забування, що викликається, якщо активність не повторюється належним чином, можна отримати ефект при часто повторюваних візерунків поїздити всю кору, щоб пропагувати свої хвилі, і рідкісні комбінації створюють ділянки місцевого поширення. Крім того, розмір цих зон не фіксуються, але помітно зростає, якщо відповідне явище починає проявлятися частіше. Ви можете знайти деякі причини в цій поведінки. Можливо, реальна кора поводиться в схожий спосіб.
Коли я змогла формувати принципи хвилі, описані вище, моделювання нашого проекту переходив на новий якісний рівень. Відчувається, як замість кам'яної осі, ми раптом отримали джеммер. У наступних частинах я покажу, як використання моделі нейромереж хвиль може пояснити багато раніше загадкових властивостей мозку.
Продовження
Література використана
Якщо щось занадто короткий, незнімний або непристойний, будь ласка, відписатися в коментарях.
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/214663/







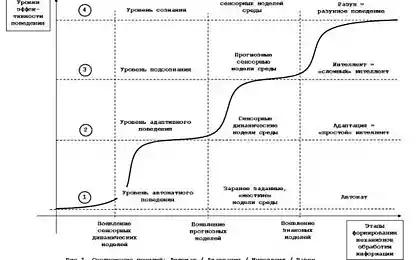











Логіка мислення. Зареєструватися 4. Підземна активність
Логіка мислення. Зареєструватися 6. Система проекції