1677
Логіка мислення. Частина 15. консолідація пам'яті

Ця серія статей описує модель хвилі мозку, що істотно відрізняється від традиційних моделей. Я дуже рекомендую, що ті, хто тільки приєднався до читання першої частини.
Давайте спробуємо уявити інформацію про мозковий мозок. З пам'яті на основі нейронів ми спочатку з'ясуємо, як багато нейронів ми працюємо. Найточніша оцінка до дати належить Сусанне Геркулано-Huzel (Frederico A.C. Azevedo, Ludmila R.B. Carvalho, Lea T. Grinberg, Хосе Marcelo Farfel, Renata E.L. Ferretti, Renata E.P. Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009). За даними кошторису, корові рахунки за 82% загальної маси мозку і 19% загальної кількості нейронів (рис. нижче). За цією оцінкою кора складається з близько 16 мільярдів нейронів (1.6x1010).
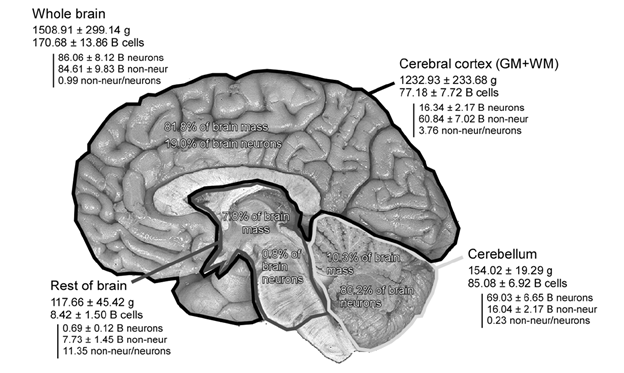
Кількість нейронів в різних частинах мозку (в мільярдах) (Frederico A.C. Azevedo, Ludmila R.B. Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata E.L. Ferretti, Renata E.P. Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009)
У моделях, які використовують синоптичну пластику, щоб пояснити характер пам'яті, пам'ять, досить простий. Типове число сипсів для нейрона 10000 (104). Якщо ми припустимо, що один сипс зберігає один біт інформації, ми отримуємо 1.6x10 x 104 = 1.6x1014 біт або приблизно 2x1013 байтів. Ось близько 20 000 гігабайтів або 20 терабайтів. Обсяг досить скромний, навіть за сьогоднішніми стандартами.
Якщо ми припустимо, що активне життя людини становить близько 80 років, з яких третина витрачається в сон, то це 80x365x(24-8)x60x60 = 1681920000 (1.7x109) секунд. Виходить, що якщо ви не враховуєте організацію пам'яті, але просто заповнюєте його протягом життя, ви можете дозволити написати швидкість 2x1013/1.7x109=1.18x104 байтів / другий або близько 10 kbytes / другий.
У нашій моделі оцінка кількості пам'яті дещо відрізняється. Оцінка сиптичного пам'яті не змінюється, те ж 20 терабайтів. Але це просто пам'ять, яка зберігає зображення детектора нейронів. Основні обсяги падають на екстрасинаптичні рецептивні кластери. Якщо один екстрасинаптичний образ утворюється з сигналів близько 100 сипсів, то можна оцінити інформаційну ємність одного елемента як 100 біт. Виходячи з оцінки, поверхня нейрона може містити близько 100 000 рецептів кластерів, ми отримуємо пам'ять 1,6х10х105х100 = 1.6 x1017 біт або 2х1016 байтів. Що таке 20 000 терабайтів або 20 терабайтів.
Припустимо, що половина цієї пам'яті витрачається на здатність кори пропагувати довжини хвиль ідентифікаторів, пам'ять події можна оцінити на 10 вихованців. Цей об'єм командує деяку повагу. На його основі швидкість запису інформації з безперервною пам'яттю може бути 1016/1.7x109 = 5.88x106 байтів / другий або близько 6 мегабайтів / другий. Якщо припустимо, що фіксація грамів відбувається кілька разів на секунду, ми можемо оцінити інформаційну спроможність одного грама в області 1 мегабайту.
Ми можемо самі зателефонувати одержувачу і узгодити зручний час і місце вручення квітів, а якщо необхідно, то збережемо сюрприз. загальність пам'яті підтверджено наявністю феноменальних людей, які дійсно “запалюють все”. Олександр Лурі, в результаті багатьох років спостережень, задокументовано один приклад такої безмежної пам'яті:
Виявилося, що пам'ять Ш. не має чітких меж, не тільки в її обсязі, але і в силі збереження слідів. Експерименти показали, що він може успішно відтворювати будь-яку довгу серію слів, наведених йому на тиждень, місяць, рік, багато років тому. Деякі з цих експериментів, які незмінно закінчилися успіхом, проводилися 15-16 років після початкової запам’ятовування серії і без попередження. З цієї нагоди Ш. сидить, закривши очі, пауза, а потім скажеш: «Так, так, це було в цій квартирі... ви сидів на столі, і я був на скелястій дошці... ви були в сірому костюмі і подивилися на мене, як це... Я бачу, що ви сказали мені...» – а потім пішов з нездатним відтворенням серії читання (Лурія, 1968).
Але неважливо, як обмежити нашу пам'ять, добре відомо, що набагато простіше і більш докладно запам'ятати останні події. Назад у 1885 році психолог Герман Ебббінгхаус отримав забуту криву (Ebbinghaus, 1885) (рис. нижче).
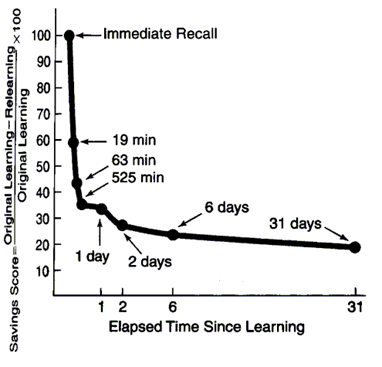
Забудьте криву
Найточніша забувка спостерігається в перші хвилини, після чого процес забування сповільнюється. Після 30 днів пам'ять надходить на стабільну фазу, де не відбувається подальше забування. У своїх експериментах Ebbinghaus використовуються набори безглуздих трилетових слів, які запам'ятовуються. Навчання дещо відрізняється від простого сприйняття, тобто забута крива для колись чув фразу або зображення буде дещо різним.
Для того, щоб пам'ятати щось більш сильно дозволяє повторювати. Ми не просто освіжаємо спогади і починаємо новий цикл забуту. Після повторення, асимпто, на яку забуває крива значно вища, ніж її попередній рівень (рис. нижче). Кожен новий повтор збільшує шанси, які після тривалого часу ми зможемо запам'ятати все відносно точно.
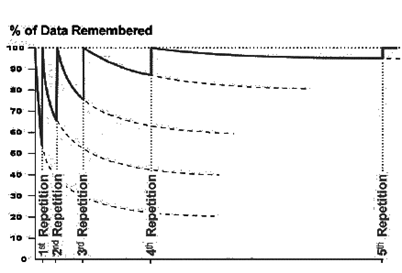 р.
р.Забудьте криву в повторі
Важливо відзначити, що повторення не обов'язково повинна бути зменшена на іншу презентацію вченого образу. Пам'ять, що точно відтворює запам'ятовуваний захід, також повторення. Це, найчастіше, ми пам'ятаємо щось, більш міцно нарізаємо в нашу пам'ять.
Традиційна інтерпретація таких властивостей пам'яті ґрунтується на ідеї передачі спогадів від короткострокової пам'яті до довгострокової пам'яті. В той же час, часто, спираючись на комп'ютерні аналогії зберігання даних, незважаючи на рух інформаційних слідів з однієї частини мозку в інший. Вважається, що hippocampus, працює як оперативна пам'ять комп'ютера, накопичує поточні спогади, а потім пересуває їх на кору, схожу на те, як комп'ютерні дані зберігаються на довгострокових носіях.
Перехід з короткострокової до довгострокової пам'яті називається консолідація пам'яті. Це об'єктивний і продуманий процес. На додаток до того, що ми можемо спостерігати зміни пам'яті з власного досвіду, є прямі докази консолідації. Так відомо, що травми голови, електричний удар, напади порушують нормальний перебіг утворення пам'яті. Вони викликають ретроградію амнезію, де загублені спогади про деякий час.
В мозку введено аналогічний ефект втрати пам'яті, коли речовини, які блокують вираз певних генів, які виробляють білки, що беруть участь у пам'яті.
Крім того, ефект ретроградованої амнесії знайомий багатьом для значної втрати пам'яті після надмірного вживання алкоголю. У деяких критичних дозах спирту так порушує процес консолідації, що деякі спогади повністю стираються, створюючи незрівнянний ляпас пам'яті. У той же час відсутні втрати здатності до поточної запам’ятовування, як при порушеннях стегна, а саме примирення вже отриманих спогадів, точніше, не переводячи їх до тривалої фази зберігання. Хворий з синдромом Корсакаоф втрачає пам'ять, як тільки його зрушення уваги. Людина, яка зробила критичну дозу, як і раніше запам'ятовує останні події, і в багатьох випадках може підтримувати пов'язану розмову. Однак вранці всі спогади, починаючи з певної точки, стираються.
Щоб пояснити ефект консолідації, не потрібно орієнтуватися на комп’ютер аналогію, пов’язані з фізичним рухом даних. У нашій моделі консолідація і його значення можна описати наступним чином. Як ми говорили раніше, екстрасинаптичні метаботропні рецептивні кластери мають ефект гістерезу. Це означає, що якщо нейрон генерує шип, то стрибає в мембранному потенціалі під ряд умов, описаних раніше, може слугувати приводом для переходу рецептивного кластера до стабільного стану. Стійкий стан - це стан, в якому кінцеві частини рецепторів виштовхуються з мембрани і рецептор стає чутливим до конкретного сигналу, або стану полоскання, в якому рецептор, навпаки, втрачає чутливість. Ця зміна відбувається з рецептивними кластерами, для яких було виконано стан втручання медіаторів з сипсів, що оточують їх, тобто які налаштовані на певне зображення.
Перехід рецептивних кластерів до цієї держави відбувається одночасно в величезній кількості нейронів, що діють в поточний час. Що означає, що цей поточний момент запам'ятовується з екстремальною почервонінням і деталями. Це призводить до того, що інші речі рівні, поточні події мають набагато більш високу ймовірність запам'ятовується у порівнянні з подіями більш віддалено.
Така гіперрозкладна пам'ять може бути викликана оперативною або короткочасною пам'яттю, але вона не повинна плутатися з поточною індукованої і хвильової активності кортичних нейронів. Діяльність нейронів формує опис того, що відбувається, при короткостроковій пам'яті зберігає фотографії таких порівняно недавніх станів. І короткочасна пам'ять не відрізняється від решти пам'яті, крім того, свіжі спогади більш детальні, оскільки вони використовують значно більшу кількість сприйнятливих кластерів, ніж інші грамоти.
Тривалість надопераційної пам'яті секунд. Після чого більшість екстрасинаптичних рецепторів повертаються до свого первісного стану, стали готові брати участь у нових діях меморанізації. Але значна частина рецептивних кластерів з тієї причини або іншої залишається в стані фіксації зображення. Вони переходять в наступну стабільну фазу, що дозволяє їм існувати далі і зберігати фрагмент своїх спогадів відповідно.
Можна припустити, що кожна пам'ять проходить через кілька етапів консолідації, втратити на кожному етапі частини її рецепторів. Найпростіший алгоритм забуту є імовірністичним. При регулюванні ймовірності рецептивного кластера, що проходить наступний етап консолідації, можна отримати необхідну кількість для кожної пам'яті за часом, передається на довгострокову пам'ять.
Консолідаційні фази супроводжуються синтезом білків, які визначають загальний перебіг всіх процесів. Процес консолідації метаботропних і іонтропних рецепторів має свої особливості, але досить схожий ідеологічно. Відразу після навчання в ядрі нейрона починається експресія негайних «земних генів». Ці гени закодують транскрипцію. Транскриптові фактори, що запускають транскрипцію генів «латого». Приблизний графік такої процедури показано на малюнку нижче.
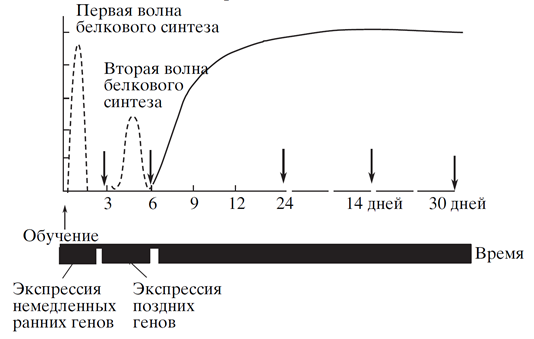
Висловлення генів «сірого» та «латого» (Є.В. Муравива, К.В. Анокін, 2006).
Є різні білки, які були показані до участі у формуванні пам'яті. Arc, CPEB, Kibra є лише кілька з них, хоча найбільш часто згадується. Пам'ятайте, що інформація залишає два види слідів на нейроні. Один пов'язаний з зміною чутливості його знімків, інші з фіксацією змін в екстрасинаптичних рецепторів. Перший впливає на властивості нейрона як детектора. Другий забезпечує можливість нейронів для поширення ID хвиль, зберігання пам'яті події, формування асоціативності, і т.д. Як і в якому послідовності відбувається регулювання білків цих процесів не простого питання і ще не має загальноприйнятої відповіді, оскільки не існує універсально визнаної моделі пам'яті. Тому в нашій моделі ми зосередимося на загальній ідеології процесів, на жаль, не ім'я можливості продемонструвати своє білкове регулювання.
Консолідація фактично встановлює таймера, який підбиває етап життя елементів пам'яті та регулює ймовірність переходу на наступну фазу. Припустимо, що повторення досвіду, або що еквівалентно запам'ятати його, змінює курс консолідації, підвищуючи шанси на сприйняття кластерів, що переходять в фазу довгострокової пам'яті. Що таке, чим частіше повторюється або частіше спогадів, тим більш значним знаком листя події. І чим вище ймовірність її відновлення з відповідним запитом інформації.
Оскільки емоційно значущі події викликають нас частіше, ніж нейтральні події, це може бути одна з причин, чому емоційно заряджені події утворюють сильніші і більш кінцеві сліди в наших спогадах.
Але спогади про досвід не так просто, як вони здаються. Психологи давно відомі явища трансформації пам'яті. Вперше показаний в 1932 році Фредерік Бартлетт (1932). Він знайшов, що якщо людина додає деталі до конкретного заходу, оригінальна пам'ять замінюється новим, а змінні деталі стають частиною цієї нової пам'яті. І людина не знає цих змін, але вважає, що все було точно так, як він запам'ятовує. Ця зміна пам'яті називається повторенням пам'яті.
Реконсолідація пам'яті добре вписується в нашу модель. Пам'ять заходу супроводжується відновленням його унікального ідентифікатора. Якщо з будь-якої причини є спотворення оригінальної пам'яті, то поточний інформаційний образ запам'ятовується таким же ідентифікатором як основна пам'ять, так як цей ідентифікатор присутній в кортиці. Таким чином, явище перетворення описане Barlett неминуче.
Є дуже цікаві експерименти, які показують, що реконсолідація може супроводжуватися не тільки трансформацією, але і епоху пам'яті. Але оскільки ці враження відносяться до поведінки, ніж до пам'яті події, ми обговорюємо їх пізніше в відповідному розділі.
Загальна ідеологія пам'яті події можна описати наступним чином. Взяти інформацію створює свіжі спогади з великою вишуканістю та деталями. Це робить їх надзвичайно доступними для швидкого доступу. Що пам'ять не триває. Займається етапами консолідації, кожен з яких зменшує кількість рецепційних кластерів, які утворюють граматики. В результаті всіх етапів консолідації, що, очевидно, триває близько місяця, занурення набувають стабільний обсяг і переходять в фазу тривалого зберігання.
Раніше ми оцінюємо порядок пам'яті події мозку на 10 вихованців. З якого методу гіпсії отримано кошторисну оцінку величини заграми 1 мегабайт. Ця оцінка може бути вірною для старих спогадів, які пройшли через всі етапи консолідації. Більшість інформації, яку ми використовуємо, є актуальною для дня. Для цієї інформації надана екстенсивна сховища. Припустимо, що така фактична пам'ять займає значну кількість пам'яті головного мозку. Якщо я був дизайнером мозку людини, я б доцільно повторити половину всієї наявної пам'яті і відібрати решту половину як старі спогади.
Під цією емпіричною припущенням, швидкість запису мозку може бути оцінена як 5x1015/(16x60x60), яка становить близько 1011 байтів або 100 гігабайтів на секунду. Звичайно, це набагато більше всієї кількості інформації, що надходить від почуттів. Але ми не говоримо про вхідний обсяг, але про докладний опис, що складається з багатьох описів, які відбуваються на кожному з кортичних зон.
У цій кошторисці початковий розмір одного грама буде близько 10 гагабайтів. Фактично процес консолідації пам'яті є поступовим зменшенням цього надзвичайно зайвого обсягу до рівня близько 1 мегабайту.
Однак, незалежно від того, що співвідношення нової і старої пам'яті фактично було, природа повинна оптимізувати її таким чином, щоб запобігти зайвому почервоніння. Це означає, що дана надзвичайно висока швидкість запису поточної інформації, може бути час переповнення всіх доступних пам'яті. Ми можемо припустити, що сонливість є державою, коли інформаційні ресурси мозку надходять на їх заповнення, а очищення пам'яті необхідна.
В принципі швидке закріплення пам'яті відбувається постійно, безперервно стискаючи обсяг грамів, мозок забезпечує простір для нових спогадів. Але ми можемо припустити, що операційна консолідація має ряд недоліків. Таким чином, у нашій моделі, детектор нейронів утворюють візерунки, які відповідають поняттям. Кожна концепція формується багатьма візерунками, розташованими в різних місцях кори. Для запуску хвилі необхідно активувати значну частину нейронів патерна. Це означає, що якщо ми використовуємо імовірнісну консолідацію, просто знімаючи деякі рецептури кластерів, ми поступово знищимо всі анграми до точки, де жоден з патернів не зможе випромінювати хвилі, пов'язані з цією пам'яттю. Щоб уникнути цього, слід використовувати вибіркові консолідації.
Вибіркова консолідація передбачає, що вибрано ми визначаємо, на які візерунки залишають чіткі сліди спогадів, а на яких повністю їх видалити. Потім з однаковим об'ємом грами і зберігаючи її поширеним характером, можна уникнути низької щільності грамів на окремих візерунках.
Припустимо, що це вигідно використовувати для збереження грамів ті візерунки, які, за їх положенням на корі, найбільш точно відповідають активності, пов'язаному з загальним контекстом пам'яті. Але процедура визначення таких візерунків вимагає відтворення пам'ятних подій. Однак таке розмноження несумісне з нормальною функцією мозку, так як, приступаючи одночасно з поточним сприйняттям, воно буде сприймати мозок як галюцинація. Припустимо, що оптимальна консолідація, пов'язана з видаленням більшості накопиченої інформації протягом дня, неможлива або складна з нормальною функцією мозку. Потім виходить, що сон є точно державою, що мозок повинен виконувати такі процедури.
На додаток до акумуляції спогадів подій мозок також просуває нейрони як детектори. І ґрунтуються на змінах структури нейрорецептори. Зміни у рецептори контролюються виразними продуктами певних генів. Ген експрес-продукти є білками. Деякі з них безпосередньо беруть участь у клітинних процесах, деякі фактори, які регулюють подальше експресування гена. При зміні концентрації відповідних білків можна судити про перебіг процесів, пов’язаних з різними видами консолідації.
Ви можете побачити і оцінити концентрацію білків шляхом фарбування ділянки мозкової тканини з барвником, який вибірково реагує на необхідний білок. Схожі спостереження показали, що найбільші зміни відбуваються під час сну (Chiara Cirelli, Giulio Tononi, 1998) (Cirelli, 2002) (див. нижче).
 р.
р.Розподіл протеїну дуги в парітал корі щура через три години сну (S) і через три години спонтанної прокидки (W) (Cirelli, 2002)
 Р
РРозподіл транскриптового фактора P-CREB в коронарних регіонах паріталової кори щурів через три години сну (S) і в разі депривації сну трьома годинами (SD) (Cirelli, 2002)
Консолідація пам'яті в снях відбувається на декількох стадіях. У здоровій людини починається сон з першого етапу повільного сну, який триває 5-10 хвилин. Далі йде другий етап, який триває близько 20 хвилин. Ще 30-45 хвилин падають на періоди третього та четвертого етапу. Після цього сплячий повертається до другого етапу повільного сну, після чого відбувається перший епізод сну РЕМ, який має коротку тривалість близько 5 хвилин. Під час сну РЕМ дуже часто і періодично роблять швидкі рухи під закритими повік. Якщо ви прокидаєте сновидіння в цей час, то в 90% випадків можна почути історію яскравої мрії. Ця послідовність називається циклом. Перший цикл має тривалість 90-100 хвилин. Потім цикли повторюються, при цьому частка повільного сну зменшується і пропорція сну РЕМ поступово збільшується, останній епізод якого в деяких випадках може досягати 1 годину. В середньому, з повним здоровим соном, є п'ять завершених циклів.
Етапи сну можна пояснити необхідністю обробки інформації різних типів. Алгоритми консолідації пам'яті події, пам'яті хвилі, пов'язаної з поширенням ідентифікаторів, а пам'ять, пов'язана з сипучою пластичністю може істотно відрізнятися.
У наших припущеннях добре пояснюється циклічна природа сну. Повна консолідація, як кожен знає від особистого досвіду, займає близько восьми годин. Циркумстанці можуть зробити прокидку раніше. Якщо восьмий час сну складається з одного циклу, розділеного на кілька фасонів, передчасне переривання сну призведе до того, що початкові фази сну встигнуть завершити, незважаючи на те, що пізніше не почалися. Тобто деякі дані залишаються повністю необробленими, при цьому повністю вичерпається відповідним мозковим ресурсом для попереднього дня. Що це, мозок, насправді, буде неприпустимо. Кілька циклів дозволяють передчасному пробудження принаймні частково виконувати кожну процедуру консолідації і принести мозок до належного стану. Але необхідність повного закріплення не зникне, а пізніше мозок доведеться компенсувати пропущені цикли з більш тривалим соном, які зазвичай називають «згортання».
Здається, що консолідація пам'яті пов'язана з повільним соном. Як вона завершена, ці фази стають коротшими, даючи спосіб швидкого сну. Це пояснює те, що при завершенні завершення консолідації завершальний етап сну складається виключно з тривалої фази сну РЕМ.
REM сон є дуже мрія, що формує дивний фантазії тільки віддалено пов'язаний з реальним досвідом. Візерунок, зазначений для снів Сігмунд Фрейд, говорить про те, що уві сні з'являються явища, які найбільш турбують нас, важливі для нас, концентрують сутність наших трупів і досвіду (Freud, 1900). У більшості випадків мрії швидко забуті. Ми мріємо кожну ніч, але стрімкість, з якою ми забуваємо, ведемо багато вірити, що вони не сниться.
У нашій моделі стирання снів досить просто. Для цього достатньо зробити сліди hippocampal ідентифікаторів, які не зафіксували протягом тривалого часу, але підлягають швидкому розпаду. Це призведе до того, щоб забути про мрії. Але якщо, прокинувшись, ми встигли запам'ятати нещодавній сон, то виникне нова традиційна пам'ять, яка збереже події мрії.
Припустимо, що для сни, стирання слідів hippocampal ідентифікаторів не супроводжується видаленням асоціативних слідів. Тоді значення снів можна пояснити, зміцнюючи систему об’єднань для найбільш значущих і важливих концепцій для нас. Це, в свою чергу, сприяє подальшому сприйняттю світу таким чином, що ці поняття будуть максимально акцентовані в системі опису нових подій.
Ми всі знаємо, що ранок вранці, ніж увечері. Але чого суть цієї мудрості? Інколи ранкові уявлення пояснюються тим, що мозок вирішує денні завдання вночі. Таке пояснення підкаже, по-перше, тим, що нічна активність мозку, записана в багатьох дослідженнях, сильно нагадує активність денного досвіду, а, по-друге, суперечка снів з тими темами, які порушують нас в реальності. Але доцільніше припустити, що мозок не вирішує завдання дня на ніч, це працює для оптимізації системи концепцій і спогадів. Вранці ця оптимізація значно полегшує пошук роз’яснень для непередбачених питань. Щоб запитати правильне питання, потрібно знати більшість відповіді. У роботі, що мозок на ніч, є приготуванням найбільш важливої частини відповіді – вміння підходити до питання з «правою» набором внутрішніх концепцій і позбутися зайвої деталі зайвих спогадів. Що таке, якщо все говорить про мрії, здається занадто вагою вам, то спробуйте повернутися до неї завтра.
Немає причин, щоб пошкодити час, проведений в сонці. Сонце – чудова можливість «забудити розумний» без зусиль. Єдине, що все ще необхідно для перевантаження голови достатньою кількістю свіжої інформації. Більш того, ця інформація може бути не тільки новими фактами, але й старими знаннями, що запам'ятовуються або думали. Всі наші думки залишають слід свіжих спогадів на корі. Уві сні з'єднуються нові відбиття і старі враження, що дозволяє більш глибоким узагальненням. Це означає, що щоденно відбиття на питання має сенс, навіть якщо вони не призводять до негайної відповіді. Ці відбиття створюють свіжі спогади, пов’язані з темою, ми зацікавлені в тому, що вночі подарує мозку можливість працювати над оптимізацією всієї системи наших концептів. Ви можете пам'ятати, що молоді діти, в яких організація кори найбільш активний, в цілому, більшість витрачених в сонці часу.
Ідеї консолідації можуть пояснити не тільки сон немовлят, але і консерватизм літніх людей. За консерватизмом ми – відхилення нових ідей та навичок, за умови збереження нормальної пам’яті подій. У традиційних підходах це викликає певний протиріччя. Здавалося б, що можливість запам'ятати нові факти повинні надати можливість дізнатися нові принципи та дізнатися нові навички. Але все потрапляє в місце, коли ви пам'ятаєте, що є організація нейронів детектора, що формує сукупність концепцій головного мозку, і є пам'ять події, яка описана і зберігається на основі цих концепцій. Щоденне консолідування пам'яті події, здається, більш ніж половина наявного обсягу. Це означає, що незалежно від віку, пам'ять події може бути сильною, якщо це не заважає органічним порушенням. Але з детектором нейронів більш складний. Здається, що з віком кількість нейронів, які визнали свою спеціалізацію, стає все більш і більше, і для формування нових концепцій є менше і менше можливостей. А потім ми приїжджаємо до ситуації, що ми можемо пам'ятати все, але тільки в поняттях, які ми маємо. Але ми вже можемо сформувати нові концепти з утрудненням або неможливістю. Відповідно, все принципово нові причини труднощів сприйняття і природного відторгнення, яке називається сенильним консерватизмом.
Література використана
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Частина 5. Брайн хвилі
Частина 6. Система проекції
Частина 7. Інтерфейс користувача
Частина 8. Ізоляція чинників в хвилях мереж
Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Частина 10. Просторова самоорганізація
Частина 11. Динаміка нейромереж. Асоціативність
Частина 12. Сліди пам'яті
Частина 13. Асоціативна пам'ять
Частина 14. hippocampus
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/216633/