1902
Логіка мислення. Зареєструватися 12. Сліди пам'яті
Ця серія статей описує модель хвилі мозку, що істотно відрізняється від традиційних моделей. Я дуже рекомендую, що ті, хто тільки приєднався до читання першої частини.
Енграми – це зміни, які відбуваються в мозку в момент запам’ятовування. В інших словах є слід пам'яті. Це цілком природно, що розуміння природи грамів сприймається всіма дослідниками як ключове завдання з вивчення природи мислення.
Що таке труднощі? Якщо ви приймаєте звичайну книгу або зовнішній комп'ютер, то як можна назвати пам'ять. Інформація про магазин. Але недостатньо зберігати. Для того, щоб отримати інформацію, необхідно, щоб дізнатися його і дізнатися, як його керувати. І ось виходить, що дуже форма зберігання інформації тісно пов'язана з принципами її обробки. Одна річ визначає інші способи.
Пам'ять людини – це не просто можливість зберігати найрізноманітніші зображення, але й інструмент, який дозволяє швидко знайти і відтворювати відповідну пам'ять. У той же час, крім асоціативного доступу до до довільних фрагментів нашої пам'яті, ми здатні зв'язати спогади в хронологічні ланцюжки, відтворювати не єдиний образ, але послідовність подій.
В рамках дослідження кортильної функції в Wilder Graves Penfield отримав добре збережене визнання за свій внесок. Займається в лікуванні епілепсії, розробляв техніку проведення відкритих бруньок, під час яких використовується електрична стимуляція, що дозволяє уточнити епілептичну спрямованість. Захоплюючи електрод в різних частинах мозку, Пенфілд записав відповідь свідомих пацієнтів. Наведено детальне розуміння функціональної організації кори головного мозку (Пенфілд, 1950 р.). Стимулювання деяких зон, переважно часових лобів, викликається яскравими спогадами, в яких минулі події наповнюються великою деталью. Рестимуляція тих же місць викликає однакові спогади.
Чітке локалізування багатьох функцій в корі, виявлених панфілом, встановлює пошук тих же чітко локалізованих слідів пам'яті. Крім того, присутність комп'ютерів і, відповідно, ідеї про те, як фізичний носій комп'ютерної інформації організовано стимулювання пошуку чогось подібного в структурах мозку.
У 1969 році Джеррі Левін сказав: «Якщо мозок людини складається з спеціалізованих нейронів, і вони кодують унікальні властивості різних об'єктів, то, в принципі, десь в мозку необхідно бути нейрон, з яким ми визнаємо і пам'ятаємо наш бабуся». фраза: рірѕрѕсѓс‚р°с‚р°с‚р°с‚р°с‚р°с‚р°с‚р°с‚р°с‚р°с‡р°с‚р°с‚р°с‚р°с‚р°с‚р°с‚р°с‚рμс‚рμс‚р°с‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμс‚рμрμрμс‚рμрμс‚с‚с‚с‚с‚сђсђсђсђсђсђсђсђсђс‚сђсђсђсђс‚с‚сђсђс‚сђсђсђсђсђсђсђсђсђсђсђсђсђсђрѕсђсђсђсђсђсђсђсђсђсђрѕсђрѕсђсђр Крім того, були виявлені прямі експериментальні докази. Нейрони були знайдені для відповіді на певні зображення, такі як чітко розпізнавання певної особи або певного явища. Однак, з більш детальними дослідженнями, виявилося, що виявлений «спеціалізовані» нейрони відповідають не тільки одному, але до груп в розумінні близьких зображень. Отже, виявилося, що нейрон, який відреагував на Дженніфер Аністон, також реагував на Ліса Кудру, який зірвав з Аністоном в телевізійних серіалах Друзі, а нейрон, який визнав Лука Скауалькера, визнаний Майстер Йода (Р. Кирог, К. Коч, І. Фред, 2013).
У першій половині ХХ ст. Карл Лашлі зробив дуже цікаві експерименти на локалізації пам'яті. Спочатку він навчив щурів, щоб знайти вихід в Мазею, а потім видалити різні частини мозку з них і відправити їх назад в Мазе. Так він намагався знайти частину мозку, яка відповідає за пам'ять придбаної майстерності. Але виявилося, що пам'ять в одному або іншому місці завжди збереглася, незважаючи на те, що значне порушення двигуна. Ці експерименти надихнули Карла Прибрама для формування широковідомої та популярної теорії голографічної пам'яті (Pribram, 1971).
Засади голографії, як і сам термін, були винайдені в 1947 році Денешом Габором, які отримали Нобелівську премію з фізики в 1971 році. Суть голографії. Якщо у вас є джерело світла з стабільною частотою, то ділите його за допомогою напівпрозорого дзеркала на два, ви отримуєте два когерентні світлові протоки. Один потік може бути спрямований на об'єкт, а другий на фотографічну пластину.
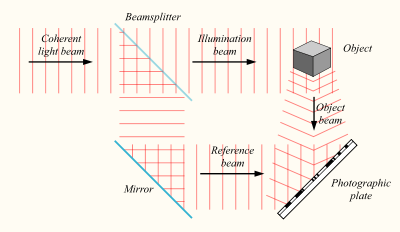
Створення голограми
В результаті, коли світло відображеного світла досягає фотопластичної пластини, він створить інтерференцію з підсвічуванням потоку пластин.
Візерунок інтерференції, відбитий на фотопластичній пластині, зберігатиме інформацію не тільки про амплітуду, а й про фази характеристики світлового поля, відображеного об'єктом. Тепер, якщо ви освітлюєте раніше виставлену пластину, буде відновлено оригінальний світловий потік, і ми побачимо пам'ятний об'єкт у всьому об'ємі.
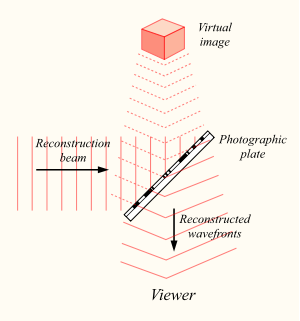
Відновлення голограми
Голограма має кілька дивовижних властивостей. По-перше, світловий потік зберігає об'єм, тобто дивиться на об'єкт phantom з різних кутів, можна побачити його з різних сторін. По-друге, кожен розділ голограми містить інформацію про весь світле поле. Отже, якщо ми вирізати голограму навпіл, ми побачимо лише половину об'єкта. Але якщо ми нахилимо нашу голову, то за межі решти голограми ми можемо бачити другу «вирізати» частину. Так, чим менший фрагмент голограми, тим нижче його постанови. Але навіть через невелику площу можна, як через брелок, подивіться весь образ. Цікаво, якщо на голограмі з’являється скеля, то через неї можна буде побачити інші об’єкти, які захоплюють їх з масгніфікацією.
З точки зору пам'яті Прибрам заявив: «Суть голографічної концепції полягає в тому, що зображення відновлені при їх представленнях у вигляді розподілених інформаційних систем належним чином активуються».
Визнання голографічних властивостей пам’яті можна знайти в двох контекстах. З одного боку, зателефонуйте голографію пам'яті підкреслює свою поширену природу і вміння реконструювати зображення за допомогою тільки дробу нейронів, схожих на що відбувається з фрагментами голограми. З іншого боку, передбачається, що пам'ять, яка має гомограмно-подібні властивості, спирається на ті ж фізичні принципи. Останнє означає, що з голографії ґрунтується на фіксуванні інтерференційного шаблону світлових потоків, пам'яті, очевидно, якось використовує шаблон інтерференції, що призводить до імпульсного кодування інформації. Відомі ритми мозку, і де є вібраційні хвилі, і, отже, їх втручання неминуче. Таким чином, фізична аналогія виглядає досить доречно і привабливо.
Але факт втручання тонка, незначна зміна частоти або фази сигналів повинна повністю змінити її шаблон. Однак мозок успішно працює з суттєвою варіацією в своїх ритмах. Крім того, спроби перешкоджати поширенню електричної активності шляхом виявлення її розділів і розміщення мики в місцях надрізів, нанесення смужок золотої фольги для створення контуру, створення епілептичного вогнища через ін'єкції алюмінієвої пасти не турбують мозок занадто патологічно (Pribram, 1971).
Висловлення пам'яті, неможливо ігнорувати відомі факти про підключення між пам'яттю і hippocampus. У 1953 р. пацієнт назвав Г.М. (Генрі Молісон) був видалений хірургом (В. Сковіль, Б. Мільнер, 1957). Це була ризикована спроба вилікувати тяжку епілепсію. Відомий, що видалення hippocampus одного з півкулів дійсно допомагає з цим захворюванням. З огляду на виняткову міцність епілепсії в Х.М., лікар прибрав hippocampus з обох сторін. В результаті Г.М. повністю втратив здатність запам'ятати все. Він пам'ятав, що сталося йому перед роботою, але все нове гасло його розуму, як тільки його увага зрушила.

Генрі Молісон
H.M. було багато досліджено. У ході цих досліджень проводилися численні різні експерименти. Один з них був особливо цікавим. Хворий попросив по колу п'ятикутну зірку, подивившись на неї в дзеркалі. Це не дуже проста задача, викликаючи труднощі при відсутності належної майстерності. В першу чергу він бачив. Але цікаво, кожен раз завдання дав йому легше і простіше. Він сам, в повторних експериментах, зазначив, що він очікуваний, щоб бути набагато складніше.
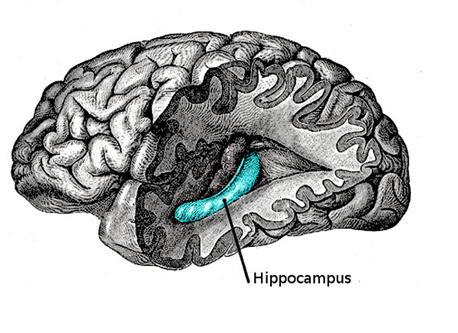
hippocampus однієї півкулі
Крім того, виявилося, що певна пам'ять подій ще властива Г.М. Наприклад, він знав про асоціацію Кеннеді, хоча це сталося після видалення його стегна.
З цих фактів було укладено, що є принаймні два різних типи пам'яті. Один тип відповідає за фіксацію конкретних спогадів, а інший несе відповідальність за отримання певного узагальненого досвіду, який виражається знаннями поширених фактів або придбанням певних навичок.
Справа Г.М. досить унікальна. В інших ситуаціях, пов'язаних з видаленням hippocampus, де не було повного двостороннього пошкодження, як в H.M., порушення пам'яті був або менш виражений або відсутній (W. Scoviille, B. Milner, 1957).
Давайте спробуємо порівнювати все, що описано в нашій моделі. Ми показали, що стійкі повторювані явища утворюють візерунки нейронів детектора. Ці візерунки здатні розпізнати їх характерне поєднання особливостей, і додавати нові ідентифікатори до хвилі. Ми показали, як функції можна змінити за допомогою ідентифікатора концепції. Це можна порівняти з пам'яттю узагальненого досвіду.
Але це узагальнена пам'ять не дозволяє відтворити конкретні події. Якщо ж саме явище повторюється в різних ситуаціях, ми в нашій нейромережі просто отримуємо асоціативні зв'язки між поняттям, що відповідає феномену і поняттям, що описує ці обставини. Використовуючи цю асоціативність, ви можете створити абстрактний опис, що складається з концепцій, які зустрічаються разом. Завдання пам'яті події не відтворювати деякі абстрактні зображення, але відтворити раніше пам'ятну ситуацію, що описує конкретну акцію з усіма його унікальними характеристиками.
Складність полягає в тому, що немає ніде в нашій моделі, де локалізується повний і вичерпний опис того, що відбувається. Повний опис складається з багатьох описів, активних на окремих зонах кори. Кожна з зон має опис хвилі з точки зору, які характерні для цієї конкретної частини мозку. І навіть якщо ми якось пам'ятаємо, що відбувається в кожній з зон окремо, ці описи все ще повинні бути пов'язані між собою, щоб виникає цілісний образ.
Схожі ситуації виникають, коли у нас є топографічна проекція та нейрони з місцевими рецептивними полями. Припустимо, у нас є нейромережа, що складається з двох плоских шарів (рис. нижче). Припустимо, що стан нейронів першого шару утворює певну картину. Цей образ передається через проекційні волокна на другий шар. Нейрони другого шару мають синоптичні з'єднання з тими волокнами, які потрапляють в межі їх рецептивних полів. Таким чином, кожен з нейронів другого шару бачить лише невеликий фрагмент оригінального зображення першого шару.
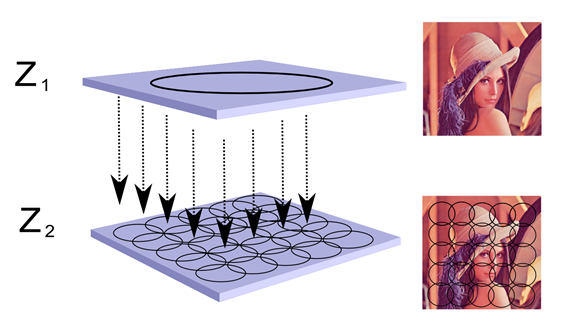
Топографічна проекція зображення на локальних рецепційних полях
На другому шарі зображено явний спосіб пам'ятати картину. Необхідно вибрати набір нейронів так, щоб їх рецептивні поля повністю закривали проектне зображення. Пам'ятайте фрагмент зображення на кожному з нейронів. І за пам'ять, щоб стати підключеною, позначте всі ці нейрони з загальним маркером, що вказує на те, що вони належать до того ж набору.
Ця пам'ять дуже проста, але надзвичайно відпрацьована в кількості залучених нейронів. Кожен новий образ вимагає нового розподіленого набору елементів пам'яті.
Ви можете отримати заощадження, якщо виявиться, що деякі загальні фрагменти повторюються в різних зображеннях, то ви не можете змусити новий нейрон пам'яті такого фрагмента, але використовувати наявний нейрон просто, додавши йому інший маркер, тепер з нової картини.
Ми приїжджаємо до базової ідеї розподіленої пам'яті. Підпишіться на нашу розсилку.
Ми проведемо різні зображення на першій зоні та проводимо їх на другу зону. Якщо ми робимо рецептивні поля нейронів досить мало, то кількість унікальних зображень в кожній місцевості не буде настільки великим. Ми можемо зробити розмір рецептивного поля так, щоб всі унікальні локальні варіанти візуалізації можуть входити в область грубо розмір нейронного рецептивного поля.
Створення просторових регіонів, що містять детектор нейронів. Зробіть кожен регіон містить детектори всіх можливих унікальних зображень і які такі регіони охоплюють весь простір другої зони. Для цього ми можемо використовувати принципи, описані вище для визначення наборів факторів.
Завдання детекторів полягає в тому, щоб порівняти зображення, подані на їх рецептивні поля з зображеннями, характерними для них. Для цього порівняння зображень можна скористатися конволюцією на рецептивному полі R:
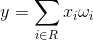
Відповідність нейрона буде більш високою, ніж новий образ охоплює зображення, що запам'ятовується. Якщо ми не зацікавлені в ступені покриття, але рівень збігу знімків, то ми можемо використовувати кореляцію зображень, що не більше нормалізованого зволоження:
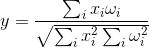
До речі, однакове значення є козином кута, утвореного вектором зображення і вектором ваги:
1 999 р.
В результаті в кожній локальній групі детекторів, детекторних нейронів, які максимально точно описують фрагмент локального фрагмента, будуть запущені, коли представлений новий образ.
Тепер давайте це: генеруйте свій унікальний тег ідентифікації для кожного нового образу та тегу активних нейронних детекторів. Ми отримаємо, що кожна презентація образу супроводжується появою на другу зону кори, яка є описом цього образу за допомогою доступних функцій другої зони. Створення унікального ідентифікатора та маркування активних нейронів детекторів із пам'яттю конкретного заходу.
Якщо ми вибираємо один з маркерів, знайдемо нейрони детектора, які містять його, і реконструювати локальні зображення, які характеризують їх, ми отримуємо відновлення оригінального зображення.
Щоб запам'ятати і відтворювати безліч різних зображень, детектор нейронів повинен мати постійні сиптичні ваги і мати можливість зберігати стільки маркерів, як вони повинні пам'ятати.
Покажіть роботу розподіленої пам'яті в простому прикладі. Припустимо, що ми виробляємо контурні зображення різних геометричних форм на верхній зоні (рис. нижче).

Подано зображення
Ми навчимо нижню зону для виділення різних факторів методом декорування. Основними зображеннями, які з'являються в кожному невеликому сприйнятливому полі, є лінії під різними кутами. Тут будуть інші зображення, наприклад, перетини та кути, характерні для геометричних форм. Але лінії домінують, тобто зустрічаються частіше. Це означає, що вони виділяються в першу чергу як чинники. Реальний результат такої підготовки показано на малюнку нижче.
 Р
РФрагмент поля факторів, виділених з контурних зображень
Ви можете побачити, що є багато вертикальних і горизонтальних ліній, які відрізняються їх положенням на приймальному полі. Це не дивно, так як навіть невеликий зсув створює новий фактор, який не перекриття його паралельними близнюками. На жаль, ми трохи ускладнюємо нашу мережу, щоб поблизу паралельних близнюків об'єдналися в один фактор. Далі припустимо, що в невеликих ділянках виявлено фактори, як показано на малюнку нижче, з певною дискретністю описують всі можливі напрямки.
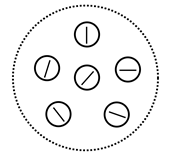
Фактори в невеликій області, що відповідають різним напрямам з дискретністю години
Після чого результат вивчення всієї коретичної зони може бути умовно зображений наступним чином:

Кондиціонерський результат підготовки коретичних зон. Для чіткості нейрони не розміщуються на регулярній сітці.
Тепер ми збираємось поставити квадратний образ на підготовлену кору. Нейрони, які дивляться характерний стимул у їх рецептивному полі, активуються (рис. нижче).
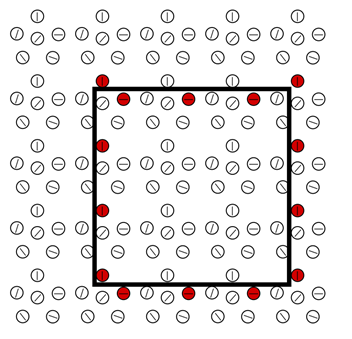
Реакція підготовленої зони кори до квадратного зображення
Тепер ми створюємо випадковий унікальний номер – ідентифікатор пам’яті. Для простоти ми не будемо використовувати наші хвилі мережі зараз, ми обмежимо себе припущенням, що кожен нейрон може зберігатися, крім сиптичних вагань, набір ідентифікаторів, тобто певного великого масиву порушень чисел. Зробіть всі активні нейрони запам'ятовують новостворений ідентифікатор у своїх наборах. Насправді, з цією дією ми зафіксуємо пам'ять квадрата, яку ми побачили.
Подаючи нові зображення, ми генеруємо унікальний ідентифікатор для кожного з них і додаємо його до нейронів, які відповіли на поточний образ. Тепер, щоб пам'ятати щось, потрібно взяти відповідний ідентифікатор, активувати всі нейрони, які містять його, а потім відновити зображення, характерні для цих нейронів. Природно, багатше і очищає систему опису, більш точно відновлене зображення збігається з оригінальним. Але навіть на дуже грубих моделях, таких як мережа вище, ви можете отримати досить помітні результати відновлення (рис. нижче).
 Р
РОригінальний образ і пам'ять, реконструйований факторами моделі «грубо»
Тепер ми можемо сформувати наші припущення щодо того, як влаштовується пам'ять події реального мозку.
Вивчення різних зон кори призводить до утворення закономірностей нейронів детектора, які здатні реагувати на зображення, характерні для цих зон. У серці цього навчання синоптична пластичність. Цей тренінг не записує конкретні події, але тільки висвітлює узагальнені поняття. При виникненні «нерухомості» його синоптичних ваг не можна використовувати для суддівських спогадів, пов’язаних з бабусям. Синапсові ваги не описують конкретні події, але зображення ознак, характерних для розпізнавання бабуся.
Картина опису, що відбуваються на кожному з кортичних зон, є подвійним. Це як картина заподіяної активності нейронів-детекторів, так і для її власних випромінювальних шаблонів для кори та хвиль, які надійшли через проекційну систему.
Роль hippocampus полягає в тому, щоб створити унікальний ідентифікатор для кожної пам'яті і додати його в загальний малюнок хвилі. В результаті на кожній зоні кори в хвильовій ідентифікаторі, крім переліку ознак, що описують поточну подію, з'явиться унікальний додаток від hippocampus, що дозволить відрізнити хвильові описи подібних подій.
Детективні нейрони в державі провокованої активності зафіксують поточний ідентифікатор хвилі на метаботропних кластерах. До речі, ми вже спостерігали щось схоже при описі системи узагальненої асоціативності концепцій. До ідентифікатора додано унікальний компонент hippocampus. З цією дією ми створюємо анграму, що дозволяє нам знайти всі нейрони детектора, які належать до однієї пам'яті.
Варто зазначити, що цей дизайн працює однаково незалежно від того, як коретична площа отримує інформацію. Для топографічної та хвильової проекції залишаються незмінні принципи запам’ятовування.
Цей дизайн пам'яті має всі голографічні властивості, необхідні для пам'яті. Нейрони-детектори будь-якого детектора шаблону зберігають інформацію про весь хвильовий візерунок, який був на корі в момент запам'ятовування, який відповідає точно, як фрагмент голограм зберігає інформацію про хвильовий візерунок навколишнього простору.
Фіксація анограми розподіляється на всі частини мозку, які показали активність в розпізнаванні чого відбувається. Це означає, що пам'ять не прив'язана до одного або декількох нейронів і не має специфічної локалізації. Вилучення будь-якої частини кори, як в експериментах Лашлі, не знищує цілу граму, але тільки переважає її описувати в умовах, які були видалені.
Це стає чітким характером нейронів, які відповідають Дженніферу Аністону або магістру Йоді. Це не нейрони пам'яті, вони тільки детектори нейронів, пов'язаних з поняттями, пов'язані з відповідними плівками.
Можна пояснити характер яскравих поглядів, які виникають при стимуляції мозку електродом. Електрод збуджує випадковий візерунок нейронів, дотик. Якщо виявиться, що цей візерунок схожий на фрагмент хвилі будь-якого відомого ідентифікатора кори, то він викликає відповідну хвилю, яка будує решту інформаційної картини мозку. Повторний електричний імпульс в вже ін'єкційний електрод викликає однакове бачення, оскільки він створює той самий малюнок активності. Але природа бачення не пов'язана з місцем, де впаде електрод. Не є поняття, які насправді мають локалізацію, яка активується, але хвиля, яка, в принципі, може виникнути в будь-якій точці. Просто на сайті електродів, візерунок цієї хвилі збігається з формою голки. Для хвильової моделі кори все це досить натуральне, але це викликає плутанину серед тих, хто шукає «грибні нейрони».
Наша концепція пам’яті добре пояснює особливості пацієнта Х.М. Оскільки hippocampus необхідний для створення унікального ідентифікатора, не дивно, що його відсутність не вдалося створити нові спогади, не порушуючи спогадів вже в існування. Де вже було призначено ідентифікатор, hippocampus не потрібно для подальших процедур інформації. З моменту утворення сповіщувачів нейронів і детекторів шаблонів не прив'язана до стегна, збереження здатності до процесуального навчання і формування узагальненої пам'яті також пояснюється.
Ще раз, коли описуємо пам'ять подій, ми не використовували синоптизацію як інструмент пам'яті. Пластика сипсів - це механізм формування закономірностей нейронних детекторів. Це, сліди конкретних подій не можна знайти безпосередньо на сипсах нейронів, хоча зображення, описані сиптичними вагами, завжди нагадують щось від попереднього досвіду. Ми приїжджаємо до необхідності окремих механізмів навчання та пам'яті подій. Виходячи з цього, наша модель виготовила два види грамів. Один тип - модифікація сиптичних ваг, що дозволяє виділити особливості на основі яких створюються всі наступні описи. Другим типом є формування екстрасинаптичних метаботропних рецептивних кластерів, які об’єднують багато нейронів, залучених до опису конкретної події. Другий тип анграми неможливий без першого. Це означає, що для того, щоб сформувати спогади про будь-які події в їх повноті, необхідно бути чинники, які дозволяють такий опис.
Інформація описана ієрархічно мозком, висвітлюючи все більш абстрактні функції від рівня до рівня. Коли ми говоримо про збирання події, ми зазвичай не означає фотографічну пам'ять нижнього рівня, але досить фіксацію більш абстрактного опису, який в процесі відновлення може призвести до реконструкції оригінальної фотографії. Але для такого опису можна, необхідно, щоб утворилися відповідні фактори. Здавалося б, що це те, чому у нас немає ранніх дитячих спогадів. У віці відхилення пам'яті ми просто не встигаємо описати події.
Література використана
Продовження
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Частина 5. Брайн хвилі
Частина 6. Система проекції
Частина 7. Інтерфейс користувача
Частина 8. Ізоляція чинників в хвилях мереж
Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Частина 10. Просторова самоорганізація
Частина 11. Динаміка нейромереж. Асоціативність
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/216263/





















Логіка мислення. Зареєструватися 11. Динамічні нейромережі. Асоціативність
Логіка мислення. Зареєструватися 13. Асоціативна пам'ять