2157
Логіка мислення. Зареєструватися 13. Асоціативна пам'ять
 Габаритний зображення
Габаритний зображенняЦя серія статей описує модель хвилі мозку, що істотно відрізняється від традиційних моделей. Я дуже рекомендую, що ті, хто тільки приєднався до читання першої частини.
У попередній частині ми показали, що розподілена пам'ять може виглядати. Основна ідея полягає в тому, що загальний ідентифікатор хвилі може об'єднати нейрони, які своєю діяльністю утворюють пам'ятну картину. Для відтворення конкретного заходу достатньо запустити відповідний ідентифікатор пам'яті на корі. Його поширення відновить той же візерунок активності, який був на корі в момент фіксації цієї пам'яті. Але велике питання, як ми отримуємо необхідний ID? Припустимо, що з набору функцій ми можемо вибрати події в описі яких присутні ці функції. Тобто має бути невралний механізм, який дозволить, відповідно до опису в певних рисах, для отримання ідентифікатора підходить для цих особливостей пам'яті.
Коли ми говорили про поширення нейронних хвиль, ми приступили від того, що нейрон зберігає на екстрасинаптичних мембранах, які є учасником. Зустрівшись з знайомою картиною, нейрон зі своїм шиком створює продовження неповторного малюнка. І ось важливо, щоб нейрон не тільки здатний розпізнати хвильовий візерунок, але це частина розкидання малюнка. У своїй пропагації може брати участь тільки частина унікальної хвилі.
Щоб не плутати в наступних аргументах, ми повторимо основні властивості хвильової моделі кори. Якщо ви позначите нейрони, які належать до однохвильового малюнка, або іншим словами, ідентифікатор, ви отримуєте щось схоже на набір точок, що показано на малюнку нижче.

Прийняття будь-якого місця кори та активації елементів ідентифікатора, ми отримуємо пропагацію хвилі з активного місця, повторюючи характерний візерунок ідентифікатора (рис. нижче).

Проходячи через кожне місце скоринки, хвиля буде «просвітити» фрагментом його унікального малюнка. Таким чином, починаючи з області 1, хвилі, досягаючи площі 2, створить унікальний візерунок, визначений ідентифікатором (рис. нижче).
 Р
РЗа унікальністю шаблону можна визначити в кожному місці кори, які ідентифікатори складають хвилю.
Якщо ми відтворили візерунок вже знайомим нам у спокійному корі в області 2, він також створить хвиля, яка, поширення на область 1, створить той самий шаблон, характерний для цього конкретного ідентифікатора.
1990Р. 2199Р.
З усього цього випливає, що для розпізнавання ідентифікатора достатньо пам'ятати в будь-якому місці кори, який візерунок створюється в цьому місці за допомогою хвилі. Це можна запам'ятати як на симпатичних западах нейрона або на екстрасинаптичну частину мембрани. Пам'ять на симпазонах призводить до відкликання активності (пульсний пакет) про визнання, запам'ятовування на метаботропних рецептивних кластерах дозволяє отримувати одиночні адгезії при з'яві знайомої хвилі.
Важко відтворити ID. Для цього потрібно активувати хоча б одну групу адгезії близько розташованих нейронів, пов’язаних з бажаним ідентифікатором. В описі зворотного виступу і введення декількох медіаторів, ми просто показали механізм, який реалізує щось схоже.
Про асоціативність концепцій, ми показали, що запит, побудований на хвилях ідентифікаторів, повертає набір ідентифікаторів концепцій, пов'язаних з поняттями, що містяться в запиту.
Щоб показати механізм перебігу спогадів від асоціативної пам'яті події, нам необхідно показати, як хвиля запиту, що складається з спеціальних концепцій, може виводити унікальні тапокам'яні ідентифікатори спогадів, придатних для цього запиту. Якщо ми можемо отримати такий набір ідентифікаторів і вибрати один з них, то, запустивши цей ідентифікатор назад через кору, ми отримаємо активність детекторних нейронів, що містять цей ідентифікатор на екстрасинаптичну мембрану, і це буде еквівалентно відновленню всієї чіткої картини необхідної пам'яті.
Нагадуємо, що про наше спрощення і схема. Далі Я окреслюв можливість нейронного механізму для ассоціативної пам'яті, не претендуючи, що це працює мозок.
Давайте розглянемо структуру реальних нейронів. Нейроне тіло, сома, має обмежену площу і не може забезпечити простір для всіх синоптичних контактів. Більшість сипсів, що відбуваються в розгалуженій структурі, називається дендритом або дендротичним деревом (рис. нижче). Кількість сипсів, розташованих на дендриті, становить 10-20 разів більше, ніж кількість сипсів на сомі.
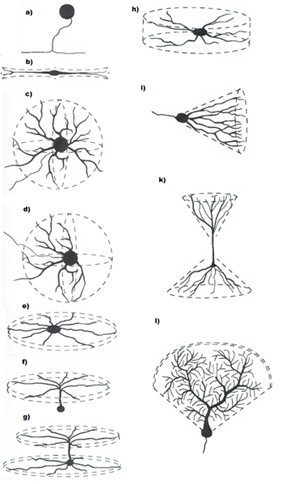
Форми дендритичних дерев (Greg Stuart, Nelson Spruston, Michael Häusser)
Виявлено, що нейрон поводиться по-різному в залежності від того, чи вступають сигнали до снайперів до одного або до різних дендритичних гілок (Шеферд Г.М., Брайтон Р.К., Miller J.P., Сегев І., Ринзель J., Rall W., 1985). Одночасний прихід імпульсів до знімків однієї гілки викликає значно сильніший нейронний відгук, ніж сигнал, що розподіляється по різних галузях.
На підставі спостережень такого роду гіпотеза народилася, що дендрицькі гілки можуть грати роль збігових детекторів (Softky, 1994). Суть його полягає в тому, що сигнали, що народжуються в віддалених гілках дендротичного дерева, щоб генерувати нейронний шик, необхідно, щоб сегменти дендротичного дерева лежать на сигналі шляху до соми.
Цей ефект був показаний для пірамідальних нейронів (Ярський Т., Алекс Роксин А., Кат В.Л., Спрустон Н., 2005), але можна припустити, що щось схоже характерне для інших типів нейронів.
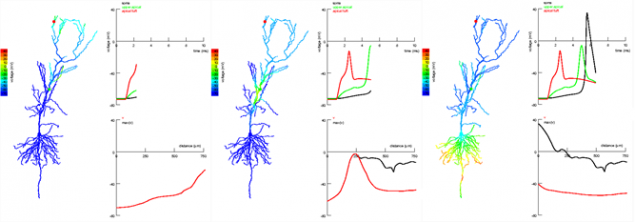 р.
р.Фази пропагації сигналів в епічних і незрівнянних сегментах пірамідальної гіпокамальної нейрона. Походження сигналу в апічний промінь (червоний точка) і поширюється на заставу Шаффера (зелена точка). Сигнал на сайті ближче до сома дендрайт не дає далекого сигналу на здачу і сприяє виникненню спіке (Ярський Т., Алекс Роксин А., Кат В.Л., Спрустон Н., 2005).
Пропагація імпульсу по дендриту супроводжується його значним нахилом. В теорії вплив віддалених (розкладних) знімків має бути значно менше впливу близьких (проксимальних) знімків. Тим не менш, механізми показали, щоб вирівняти внесок таких сипсів, що призводять до концепції «синаптичної демократії» (кліфон С. Румсі, Л. Ф. Абботт, 2006). Вирівнюючи внесок сипсів вздовж відділення дендриту дозволяє гілки розглядатися як самостійні логічні елементи, сигнали яких якось додатково обробляються нейрон. Це означає, що теоретично, змінивши конфігурацію дендриту та відповідь соми, можна отримати нейрони з різними логічними властивостями.
Наприклад, популярна концепція «ієрархічної часової пам'яті Джеффа Кукіна» використовує нейрони, які використовують автономні елементи, що працюють в режимі «або» (рис. нижче).
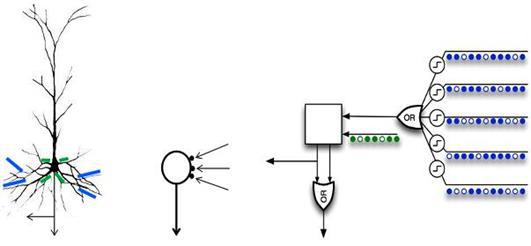
Модель нейрона з набором дендритичних логічних елементів у порівнянні з пірамідальним нейроном (Гавкінс, 2011)
Припустимо, що реальний мозок працює нейронами з значним різноманіттям властивостей.
Тепер перейдемо до опису пам'яті про асоціативну подію. Припустимо, у нас є два типи нейронів, які утворюють плоску кору. Поле рецептора цих нейронів охоплюють місцеву площу їх навколишнього середовища, де потрапляють обидва види нейронів. Ми ділимо їх на два шари, пам'ятаємо перерозподілу зв'язків.
Ми встановлюємо нейрони першого типу, щоб вони пропагували хвилю ідентифікатора інформації. Нейрони другого типу будуть змушені пропагувати лише хвилі ідентифікаторів hippocampal (рис. нижче).
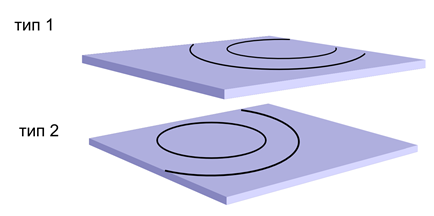
Пропагація двох незалежних хвиль на нейронах з різними типами медіатора
Для цього ми розбавляємо своїх осей і екстрасинаптичних рецепторів медіаторами, які використовуються (план нижче). Зверніть увагу, що синоптичні рецептори не важливі для визначення пропагації хвилі. Аксон тип 1 А Тип 2 Б Нейротрансміттер характеристик нейронів. Чутливість рецептора і едикція аксона
З таким завданням медіаторів і рецепторів хвилі на двох шарах не будуть впливати один на одного.
Тепер слід ускладнити другий тип нейронів, зробити їх дендротичними деревами складаються з двох типів гілок, різних додаткових синоптичних медіаторів. У моделюванні однорідні гілки можна поєднувати і в результаті два дендротичні сегменти, що діють на принципі «або» (табл нижче). Axon Тип 1 A Тип 2 1 B 2 A Нейротрансміттер Характеристики нейронів. Чутливість рецептора і едикція аксона. Нейрони другого типу мають два дендротичні сегменти з рецепторами, чутливими до медіаторів різних типів.
Припустимо, що в той час як нейрони були навчені, як пропагувати довжини хвиль ідентифікаторів, другий тип нейрона був заблокований другим сегментом, чутливим до медіатора «ален». Вся інформація, необхідна для формування хвиль в нейронах другого типу буде відкладено на екстрасинаптичну пам'ять першого сегмента.
Якщо ми зараз включають в себе другий сегмент, вони починають пам'ятати хвильові візерунки, що пропагують через нейрони першого типу. Ми припустимо, що екстрасинаптична пам'ять виникає, коли нейрон генерує шип. При цьому не записується миттєва картина активності на сипсах, але накопичується хвиля під час усього інсульту пропагації. Прискорення дозволить нам запам'ятати фрагмент першої хвилі рівня, навіть якщо хвильові фази двох рівнів на місці пам'яті не збігаються. Це, коли другий рівень нейронних шипів, він захопить картину інформаційної діяльності, навіть якщо це було деякий час назад.
Покажіть, що ми зробили таку пам'ять. Всі нейрони другого рівня, що пропагують хвилю hippocampal ідентифікатора, активні в цьому інсульті, запам'ятають на своїх другий сегментах фрагменти інформаційної хвилі, які пройшли через нейрони першого типу. Це означає, що якщо ми коли-небудь повторюємо однакову хвилю опису, то всі другий рівень нейронів, які розпізнають цю картину, попадуть. Загальна картина цих походів відтворить той же ідентифікатор hippocampal, який був в момент запам'ятовування цього образу інформації.
Так ви можете запам'ятати різні дескриптивні фотографії. Коли відповідні описи повторюються, нейрони другого рівня виготовлять повне поєднання всіх ідентифікаторів hippocampal, які потрапляють під поточний опис.
Ви можете побачити, що така пам'ять є надзвичайно відпрацьованою. Ми пам'ятаємо кожен візерунок на всіх активних нейронах другого типу. Хоча для відтворення ідентифікатора потрібно лише невелика місцева зона, яка може почати хвилю. Щоб виправити ситуацію, ми пам'ятаємо про інформаційні хвилі не на всіх активних нейронах другого типу, але тільки на тих, які потрапляють в райони з високою індукованої діяльністю нейронів першого типу. Пам'ятайте, що інформація подвійна. Це як і ідентифікатор хвилі, так і активність закономірностей індукованої активності. Хвиля подарує нам візерунок, щоб запам'ятати, і візерунки дадуть нам місця, щоб пам'ятати.
Найлегший спосіб вибору місця для використання тієї ж сфери діяльності, яку ми використовували для просторово організувати візерунки. У цій конфігурації пам'ять можна знайти наступним чином. У місцях кори, де формується інформаційний візерунок, створює візерунки індукованої діяльності, формується поле активності. Усі активні нейрони другого типу, що пропагують гіпокамальною хвилею, для якої поле активності над певним порігм, закріплені на екстрасинаптичних картинах пам'яті, описаних за допомогою інформаційної хвилі.
Подаючи інформацію про хвилювання, ми отримуємо активність нейронів другого типу, який визнає цей запит. Місцеві групи таких нейронів випромінять ідентифікатори hippocampal, пов’язані з усіма спогадами, пов’язаними з цим запитом.
Ми показали, як можуть бути захоплені і відновлені унікальні ідентифікатори пам'яті. Ми раніше показали, що якщо пам'ять таких унікальних ідентифікаторів зберігається на нейронах детектора, можна реконструювати зображення, що відповідають спогадам. У таблиці нижче показано можливість конфігурації медіаторів для такої пам'яті. Оксон тип 1 A A A 2 B тип 2 1 B B B 2 A Нейротрансміттер характеристик нейронів. Чутливість рецептора і едикція аксона. Двостороння конфігурація
Налаштування площі пам'яті через поле діяльності є досить сирим інструментом. Удосконалюється дизайн. Наприклад, через сиптичну підготовку нейронів другого типу можна створити з них регіонів, що відповідають поняттям, розташованим в цьому місці кори. За допомогою концепції датчика дендротичного матчу можна зробити активність концепції передумовою для запису та зворотного покоління ідентифікаторів пам'яті, пов'язаних з цим поняттям.
В цілому багатість інструментарію, яка забезпечує хвильову модель кори з багатосекційними нейронами, дозволяє реалізовувати досить складні і цікаві конструкції. Ми будемо обмежувати себе найпростішими схемами, які вже дали, щоб не заплутувати оповідання повністю.
Література використана
Продовження
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Частина 5. Брайн хвилі
Частина 6. Система проекції
Частина 7. Інтерфейс користувача
Частина 8. Ізоляція чинників в хвилях мереж
Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Частина 10. Просторова самоорганізація
Частина 11. Динаміка нейромереж. Асоціативність
Частина 12. Сліди пам'яті
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/216301/