3599
Логіка мислення. Зареєструватися 1. нейрон

Рік і половина тому я опублікував серію відеолекцій на Hubr з моє бачення того, як працює мозок і які можливі способи створення штучного інтелекту. З тих пір було зроблено значний прогрес. На комп'ютері було більш зрозумілим. Яким було приємно, що люди, які активно брали участь у роботі над проектом.
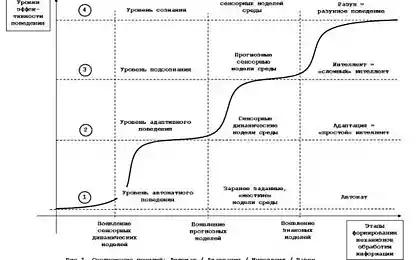
У цій серії статей планується розповісти про концепт інтелекту, який ми зараз працюємо і демонструємо деякі рішення, які принципово нові в галузі моделювання мозку. Але для оповіді бути чітким і послідовним, він міститиме не тільки опис нових ідей, але і розповідь про роботу мозку в цілому. Щось таке, особливо на початку, може здатися простим і добре відомою, але раджу не пропустити їх, так як вони значно визначають загальні докази наратив.
Загальний вигляд мозку
Нервові клітини, ака нейрони, разом з їх волокнами, які передавають сигнали, утворюють нервову систему. У хребетних основна маса нейронів зосереджена в порожнині черепа і хребта. Це називається центральною нервовою системою. Відповідно, мозок і спинний шнур відрізняють його складовими.
Спинний шнур збирає сигнали від більшості рецепторів тіла і передає їх до мозку. Через структури таламуса вони розподіляються і продаються на кори головного мозку.
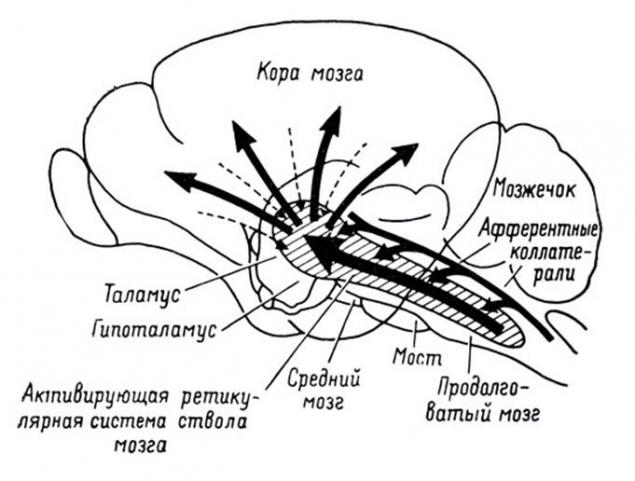
Проекція інформації на кори
Крім великих півкулів, церетель також займається інформаційною обробкою, яка, власне, є невеликим незалежним мозку. Церебелум забезпечує точну моторну майстерність і координацію всіх рухів.
Бачення, слухання та запах забезпечують мозок з потоком інформації про зовнішній світ. Кожна з компонентів цього потоку, що проходить через його шлях, також продається на скоринки. Кора - шар сірої речовини 1.3 до 4,5 мм, що робить зовнішній вигляд мозку. У зв'язку з запорами, утвореними складками, кора упакована так, щоб вона займає три рази менше площі, ніж у вигнутій формі. Загальна площа скоринки однієї півкулі становить приблизно 7000 кв.см.
В кінці всі сигнали продаються на кору. Проекція здійснюється пучками нервових волокон, які розподіляють на обмежених ділянках кори. Проведено область, на якій здійснюється або зовнішня інформація або інформація з інших частин мозку. В залежності від того, які сигнали направляють на таку зону, вона має свою спеціалізація. Відрізняється моторною зоною кори, сенсорної зони, Broca, Wernicke зон, візуальних зон, occipital lobe, всього близько сто різних зон.
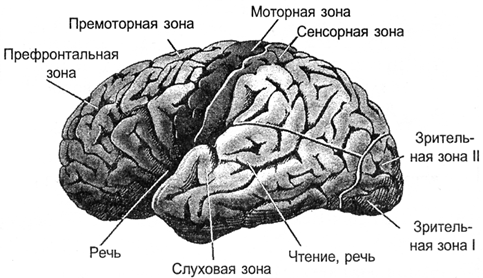 р.
р.Барк зони
У вертикальному напрямку скоринка ділиться на шість шарів. Ці шари не мають чітких меж і визначаються поширеністю певного типу клітинки. У різних зонах кори ці шари можна виражати по-різному, міцніше або слабкіше. Але, в цілому, можна сказати, що скоринка досить універсальна, і припустимо, що функціонування її різних зон підлягають однаковим принципам.

Барк шари
Відправляється сигнали на кору. Вони падають на III, IV рівень кори, де вони розподіляються на нейрони біля місця, де надійшло аферентне волокно. Більшість нейронів мають осьові з'єднання в межах своєї кори. Але деякі нейрони мають токсини, які виходять за неї. На цих еферентних волокнах сигнали йдуть або зовні мозку, наприклад, до органів виконавчої влади або продаються інші частини кори власної або іншої півкулі. В залежності від напрямку передачі сигналу, еферентні волокна діляться на:
- асоціативні волокна, які з'єднують окремі ділянки кори однієї півкулі;
- Усувні волокна, які з'єднують кору двох півсфер;
- проекційні волокна, які з'єднують кору з ядром нижніх частин центральної нервової системи.
Ви можете уявити кору головного мозку як велике полотно, розділене на окремі зони. Візерунок нейронної активності в кожній зоні зашифровано певну інформацію. Бундлеса нервових волокон, утворених осями, які виходять за межі їх коретичного поясу, утворюють систему проекційних з'єднань. На кожну зону прокладено певну інформацію. Більш того, кілька інформаційних потоків можна прибути в одну зону одночасно, яка може приходити з обох зон власної і протилежної півкулі. Кожен потік інформації схожий на своєрідну картину, намальовану активністю осей нервового пучка. Функціонування окремої зони кори – отримання набору проекцій, запам’ятовування інформації, обробки її, формування власної картини діяльності та подальшої проекції інформації, отриманої в результаті роботи цієї зони.
Важливим обсягом мозку є біла матерія. Утворюється аксонами нейронів, які створюють ці проекційні доріжки. На малюнку нижче біла матерія може спостерігатися як легка начинка між корою та внутрішніми структурами мозку.
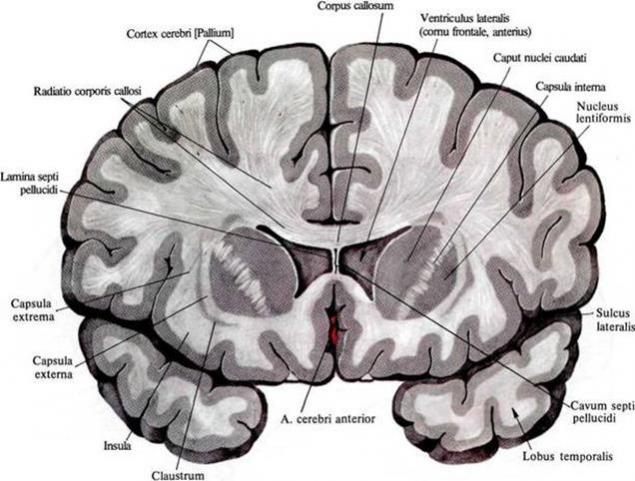
Розподіл білої матерії на передній секції мозку
За допомогою дифузних спектральних МРТ можна відслідковувати напрямок індивідуальних волокон і побудувати тривимірну модель з'єднання кортичних зон (проект підключення).
Ідея структури з'єднань добре надається кресленнями нижче (Van J. Wedeen, Douglas L. Rosene, Ruopeng Wang, Guangping Dai, Farzad Mortazavi, Patric Hagmann, Jon H. Kaas, Wen-Yih I. Tseng, 2012).
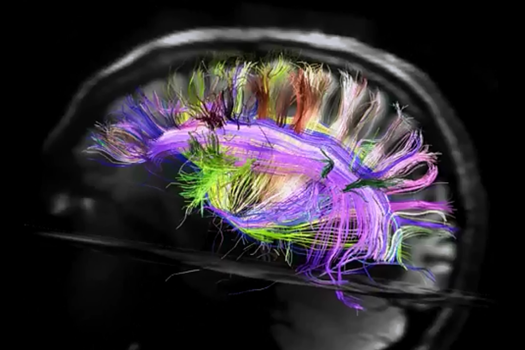
Лівий півсфера вигляд
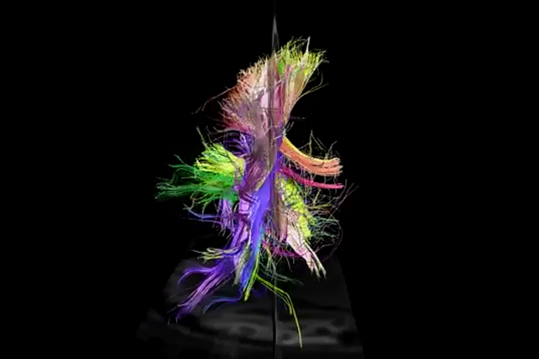 р.
р.Переглянути ззаду
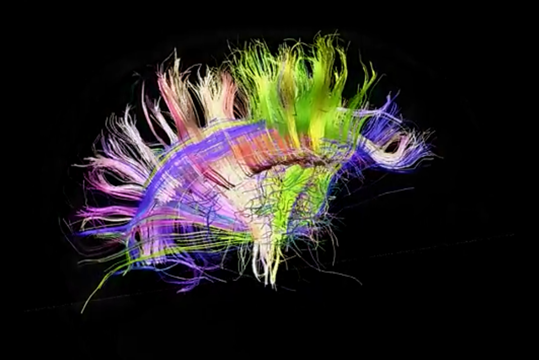
Вид з права
До речі, задній погляд чітко показує асиметрію проекційних шляхів лівої та правої півкулі. Цей асиметрія значно визначає відмінності функцій, які набувають півсфери, як вони навчаються.
нейрон
Спробон мозку – нейрон. Природно, моделювання мозку за допомогою нейромереж починається відповідь на питання, який принцип його роботи.
У серці реальної нейрони є хімічні процеси. В іншому випадку існує потенційна різниця між внутрішнім та зовнішнім середовищем нейрона – мембранним потенціалом близько 75 мілівольтів. Утворюється завдяки роботі спеціальних молекул білка, які працюють як натрієві насоси. Ці насоси, за рахунок енергії нуклеотиду ATP, приводять іони калію всередині, і і іони натрієвих натрієвих навантажень за клітину. Оскільки білок діє як ATP-ase, тобто фермент, який гідролізує ATP, він називається натрію-потазія ATP-ase. В результаті нейрон перетворюється на зарядний конденсатор з негативним зарядом всередині і позитивним зарядом зовні.
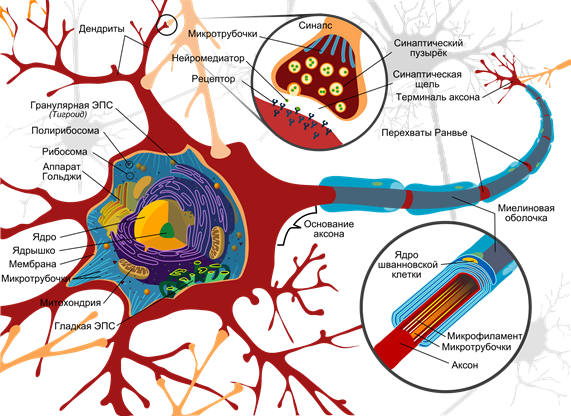
Нейронська схема (Mariana Ruiz Villarreal)
Поверхня нейрона покрита розгалуженими процесами - дендритами. Дендрити прилягають до осінніх закінчень інших нейронів. Місцезнаходження їх з'єднань називається synapses. Через сиптичну взаємодію нейрон здатний реагувати на вхідні сигнали і, за певних обставин, генерувати власний імпульс, який називається спрейкою.
За допомогою нейротрансмітаторів відбувається передача сигналів при сипсах. Коли нервовий імпульс через аксон надходить до синапсу, він випускає з спеціальних молекул нейротрансміттера бульбашок, характерних для цього сипсу. На мембрані нейрона отримання сигналу є молекули білків - рецептори. Рецептори взаємодіють з нейротрансмітаторами.
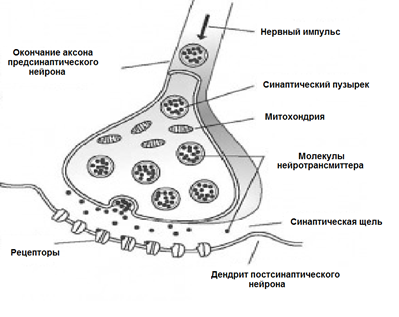
Хімічний сипс
Рецептори, які знаходяться в сиптиці, є іонотропним. Ця назва підкреслює те, що вони є іонними каналами, які можуть переміщати іони. Нейротрансмітери впливають на рецептори, щоб їх іонні канали відкриті. Відповідно, мембрана або деполяризація або гіперполяризація, в залежності від яких вражаються канали і тип сипсу. У збудливих симпазонах з'являються канали, які переходять в клітинку, а мембрана деполяризована. У гальмівних засобах, каналах, які проводять відкриті аніони, що призводить до гіперполяризації мембрани.
При певних обставинах можна змінити чутливість, яка називається синоптичним пластичністю. Це призводить до того, що синопси одного нейрона отримують різну схильність до зовнішніх сигналів.
У той же час багато сигналів наділяються на знімки нейрона. Смола гальмування натягує потенціал мембрани до накопичення заряду всередині клітини. Активувати сніпки, навпаки, спробувати розрядити нейрон (рис. нижче).
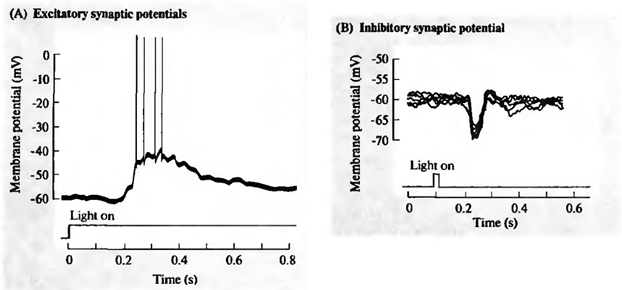
Визнання (A) та пригнічення (B) ретинальної гендерної клітини (Nichols J., Martin R., Wallas B., Fuchs P., 2003)
Коли загальна активність перевищує порогу ініціації, відбувається виділення, що називається потенціалом дії або походом. Похід є різкою деполяризації нейронної мембрани, яка генерує електричний імпульс. Процес генерації імпульсів триває близько 1 мілісекунду. У той же час, ні тривалість, ні амплітуда імпульсу залежить від того, наскільки сильні причини були (рис. нижче).
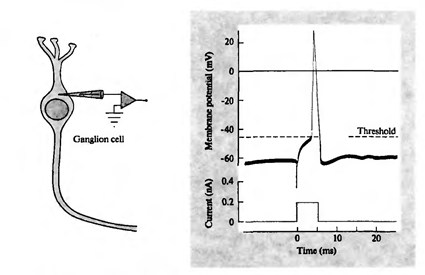
Реєстрація потенціалу дії гангліонної клітини (Nichols J., Martin R., Wallas B., Fuchs P., 2003)
Після адгезії іонні насоси забезпечують нейротрансмітерне перезаряджання і спинне затискання. Під час вогнетривкого періоду після походу нейрон не здатний генерувати нові імпульси. Тривалість даного періоду визначає максимальну частоту генерації, яка здатна нейрон.
Запобігання, що виникають в результаті діяльності при спинках, називаються індукованими. Частота відкликаних спійок полягає в тому, як добре вхідний сигнал відповідає настройку чутливості нейронних синопсів. При надходженні сигналів потрапляють на чутливі знімки, які активують нейрон, і це не заважає сигналам, що надходять до інгібіторних знімків, нейронна реакція максимальна. Зображення, описане такими сигналами, називається стимулятором, характерним для нейрона.
Звичайно, ідея того, як робота нейронів не повинна бути перенасичена. Інформація між деякими нейронами може передаватися не тільки адгезивами, але і через канали, що з'єднують їх внутрішньоклітинний вміст і передають електричним потенціалом безпосередньо. Цей пропагація називається поступовим, а сам з'єднання називається електроприводом. Дендити в залежності від відстані до тіла нейрона діляться на проксимальний (закритий) і дистальний (реот). Дендрайти можуть формувати розділи, які функціонують як напівавтономні елементи. На додаток до сипучих шляхів збудження, є додаткові синоптичні механізми, які викликають метаботропні домішки. Крім індукованої діяльності також є спонтанна активність. І, нарешті, нейрони мозку оточені гліальними клітинами, які також мають значний вплив на процеси, що відбуваються.
Довгий шлях еволюції створює безліч механізмів, які мозок використовує в своїй роботі. Деякі з них можна зрозуміти самостійно, тоді як інші можуть бути зрозумілі лише з урахуванням складних взаємодій. Тому не беремо вищеописаний опис нейрона як вичерпний. Щоб переїхати на глибокі моделі, потрібно спочатку зрозуміти «базічні» властивості нейронів.
У 1952 році Алан Ллойд Ходгкін і Андрій Гукслі розповів про електричні механізми, які визначають генерацію і передачу нервового сигналу в гігантському кальмарі (Ходгкін, 1952). Присуджено Нобелівську премію в галузі фізіології та медицини в 1963 році. Модель Hodgkin-Huxley описує поведінку нейрона системою звичайних диференціальних рівнянь. Ці рівняння відповідають автохвильовим процесом в активному середовищі. Враховуючи безліч компонентів, кожен з яких має власний біофізичний аналог в реальній клітині (рис. нижче). Іонні насоси відповідають струменню Ip. Внутрішній ліпідний шар клітинної мембрани утворює конденсатор потужністю Cm. Іонні канали сиптичних рецепторів забезпечують електричну провідність гни, яка залежить від сигналів, що передається, змінюючи час т, а загальна вартість мембранного потенціалу В. Поточний витік мембранних пори створює провідник gL. Рух іонів через іонні канали відбувається під дією електрохімічних градієнтів, які відповідають джерелам напруги з електромотивною силою En і EL.
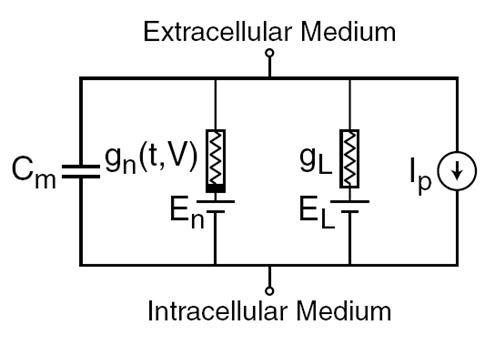
Основні компоненти моделі Hodgkin-Huxley
Природно, при створенні нейромереж, виникає бажання спрощувати модель невралу, залишаючи тільки найактуальніші властивості в ній. Найвідоміша і популярна спрощена модель Маккулох-Питтс штучна нейронна розроблена на початку 1940-х років (МакКульлоч Ю., Піттс В., 1956).
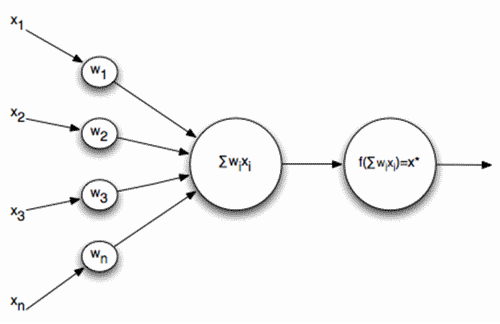
Формальний Маккулох-Питц нейрон
Заявки надсилаються до вводів такого нейрона. Ці сигнали зважуються разом. далі, функція нелінійної активації наноситься на це лінійне поєднання, наприклад, сигмоідальне. Нерідко як сигмоідальна логістика використання:
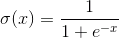
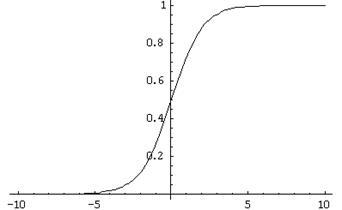
Логістична функція
У цьому випадку активність формального нейрона записана як
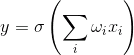
В результаті такий нейрон стає поріговим доповненням. З досить крутою функцією порогу, вихідний сигнал нейрона або 0 або 1. Зважена сума сигналу вводу і вага нейрона - це переконливість двох зображень: зображення сигналу вводу і зображення, що описано вагами нейрона. В результаті плутанини вище точності листування цих зображень. Що таке нейрон, по суті, визначає, скільки заданий сигнал схожий на зображення, записане на його сипсах. Коли значення конволюції перевищує певний рівень і порогу функція перемикається до одного, це може тлумачитися як сильну заяву нейрона, яка визнала зображення, представлене.
Реальні нейрони є дуже схожими в деякій мірі, щоб Маккулох-Питц нейрон. Амплітуда своїх спійок не залежить від того, які сигнали на сонці зумовили їх. Похід, або немає. Але реальні нейрони відповідають стимулу не одним імпульсом, але з послідовністю імпульсу. У той же час, частота імпульсів вище більш точного зображення, характерного для нейрона. Це означає, що якщо ми будуємо нейромережу з такого порогу літо, це дасть деякий вихід, але цей результат буде далеко від того, як працюють реальні нейрони. Для того, щоб принести нейромережу ближче до біологічного прототипу, нам потрібно буде імітувати операцію в динаміці, враховуючи часові параметри і відтворювати частотні властивості сигналів.
Але є ще один спосіб. Наприклад, ми можемо відрізнити узагальнену характеристику діяльності нейрона, яка відповідає частоті її імпульсів, тобто кількість спій протягом певного періоду часу. Якщо ми йдемо до цього опису, ми можемо уявити нейрон як простий лінійний підсилювач.
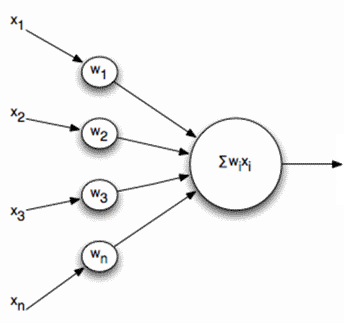
Лінійна дошка
Вивід і, відповідно, вхідні сигнали для таких нейронів більше не є діхатомним (0 або 1), але виражаються деякою кількістю скалярів. Функція активації потім написана як
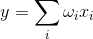
Лінійний дозатор не повинен сприйматися як щось принципово відрізняється від пульсованої нейрони, він просто дозволяє переходити до більш тривалого інтервалу часу при моделюванні або описі. І хоча опис імпульсу є більш правильним, перехід на лінійний додаток в багатьох випадках, обумовлених сильним спрощенням моделі. Більш того, деякі важливі властивості, які важко дискренувати в пульсовому нейроні, досить очевидні для лінійного додатка.
Продовження
Література використана
Олексій Редозув (2014)
Джерело: habrahabr.ru/post/214109/