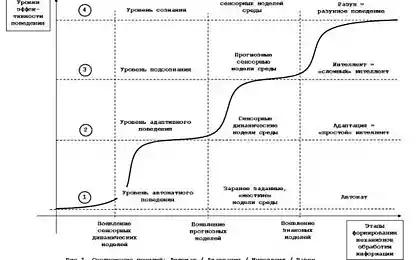
3598
思维的逻辑。第1部分神经元

一年半前,我换上哈卜尔周期 的视频讲座与我的大脑是如何工作的视野,什么是可能的方式来创造人工智能。在过去,因为时间管理向前推进。事情发生了更深入的了解,不知怎的,成功地模拟一台计算机上。什么是好的,有支持者,积极参与该项目。
在本系列文章的计划谈论上,我们正在努力,并展示一些是全新的,在大脑建模领域的决策的智力的概念。但这个故事是明确和一贯的,它将包含新的想法不仅是一个描述,也是对大脑是如何工作的一般的故事。有些事情,特别是在刚开始可能看似简单而闻名,但我劝你不要错过他们,因为他们在很大程度上决定了叙事的总体结论性意义。
大脑
的总体思路
神经细胞,它们也神经元,其纤维发射信号构成神经系统。在脊椎动物中,大部分的神经元都集中在颅腔和椎管。这就是所谓的中枢神经系统。因此,分离的脑和脊髓作为其分量。
脊髓收集来自最受体身体信号,并将其发送到大脑。通过丘脑的结构它们分布并投射到脑的大脑皮层。
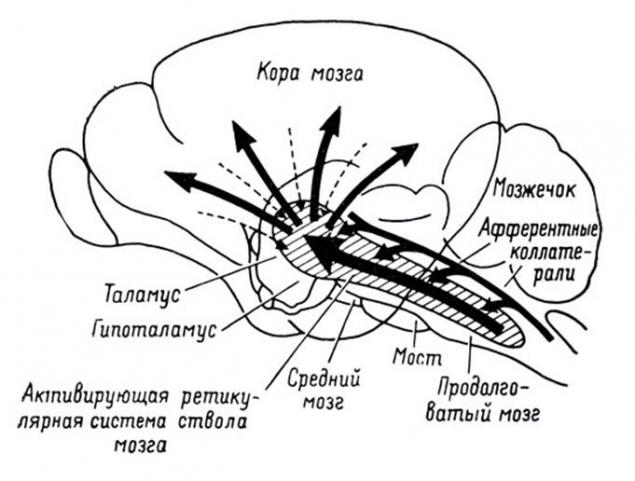
在树皮上的投影信息 i>的
除了从事信息处理大脑半球也是小脑,这,其实是一个小的独立的大脑。小脑提供了精确的运动技能和协调所有的动作。
视觉,听觉和嗅觉的大脑提供的信息对外界的流量。每个这种流的部件,通过在其路径上,并且被投影到所述外壳。树皮 - 一层厚厚的灰质从1.3至4.5毫米,构成脑的外表面。由于圈形成的褶皱,树皮填充使得它需要三倍更少的空间比在膨胀形式。一个半球皮质的总面积 - 约7000平方厘米
其结果是,所有的信号被投影到外壳。突起进行神经纤维,它们分布于皮质的有限区域的束。从脑的其它部分的图,其中被投影或外部信息,或者信息形成皮层的区域。取决于哪个信号到达这样一个区,它有其专门化。区分运动区皮层,感觉区,Broca区,韦尼克的视觉区域,枕叶,只有约一百个不同的领域。
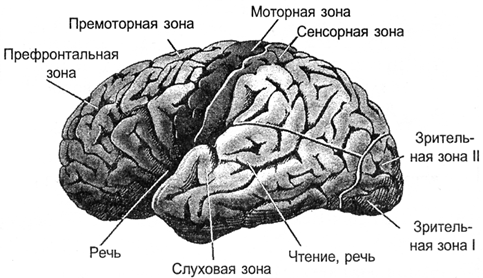
皮质 I>
在垂直方向上的皮层可以分为六个部分。这些层不具有明确的界限,并通过特定的细胞类型的流行的定义。在各种不同的皮层区这些层可以表示以不同的方式,更多或更少。但是,在一般情况下,我们可以说,在地壳是足够灵活,并假定其各个区域的功能是受相同的原则。

树皮图层 I>
由传入纤维的信号被发送到皮层。他们落在III,皮质IV水平,蔓延到附近的地方下跌传入纤维的神经元的地方。大多数神经元有皮质的区域内轴突连接。但是,一些神经元轴突有超越它。对于这些传出纤维信号或者是大脑,如执行机构,或者投射到他或另一个半球的皮层的其它部分之外。视信号传输传出纤维的方向可分为:
在联想纤维连接一个半球皮质的各个部分; 合纤维连接皮层的两个半球; 与下部的原子核连接皮质投射纤维中枢神经系统。 LI> UL>如果采取的方向垂直于皮质表面,然后注意到,神经元,其位于沿着这个方向,响应类似的刺激。这些垂直间隔组神经元,称为皮质柱。
可以想见大脑皮层的一块大布,切出的各个区域。图象每个区的神经元活动的编码的某些信息。神经纤维束通过轴突超越其皮层形成,形成的投影环节的制度。在每个区的被投影的某些信息。和一个区域可以输入多个信息流,可以配备两个他带,以及相对的半球。类似的那种画面绘制活动的信息每个流轴突的神经束。功能分开皮质 - 被提供多个突起,信息记忆,其加工,他自己的绘画活动,并进一步投影信息的形成,导致在此区域中的一个工作
。
一个显著量的大脑 - 一个白色物质。它是由神经细胞的轴突形成,创造了非常平坦的方式。在图中可以看到白色的无论多么明亮的皮质和大脑的内部结构之间的灌装。
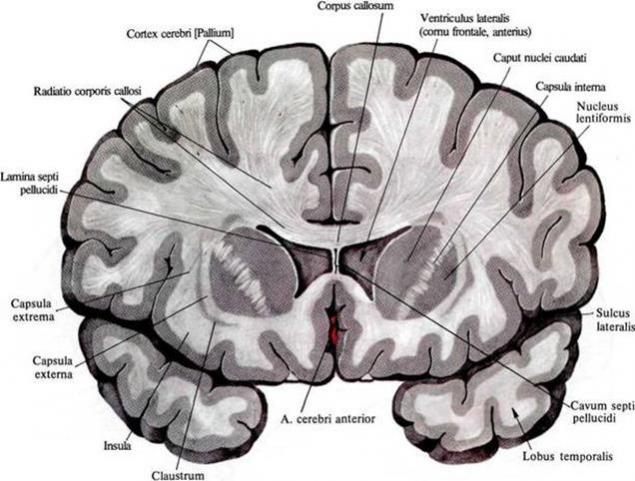
白质大脑中的 i>的
的正面部的分布
使用频谱扩散MRI能追踪单个纤维的方向,建立一个立体的连通皮质区(项目Connectomics(CONNECT))模型。
理解关系的结构是远远低于给定的数字(范J. Wedeen,道格拉斯L. Rosene,吴若鹏王,许广平戴,法扎德Mortazavi,帕特里克海格曼,乔恩·H·凯丝,文弘毅一,曾雅妮,2012)。
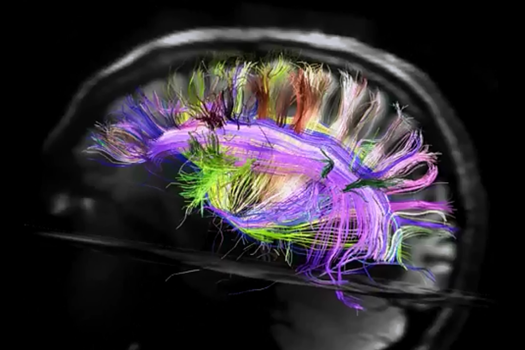
查看从左脑 I>
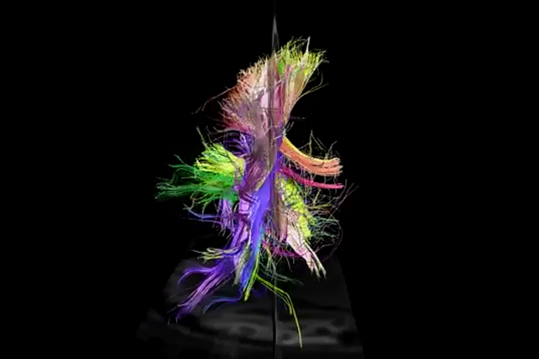
后视图 I>
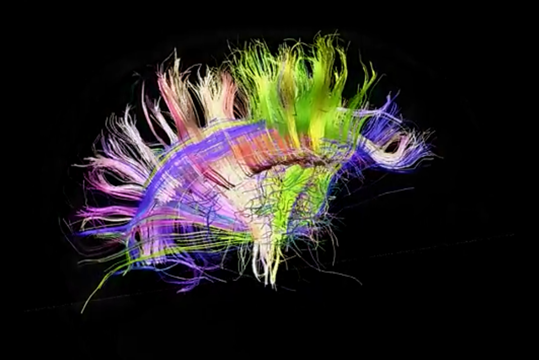
右视图 I>
顺便说一下,一个背投显然左和右半球可见不对称道。这种不对称性在很大程度上决定了那些获得半球,因为他们正在学习功能的差异。
神经元
脑基 - 神经元。自然地,利用神经网络脑的造型开始答案的问题,什么是它的工作原理。
在一个真正的神经元的心脏是化学过程。在内部和神经元的外部之间的休息状态,有一个电势差 - 约75毫伏的膜电位。它是由特定的蛋白质分子,其充当钠钾泵的操作而形成的。这些泵由于内部的能量核苷酸的ATP从动钾离子和钠离子 - 出细胞。因为蛋白质因此充当一个ATP酶,即酶水解ATP的,它被称为 - “钠 - 钾ATP酶”。其结果是,神经元被转换成一个充电的电容器与负电荷内外阳性。
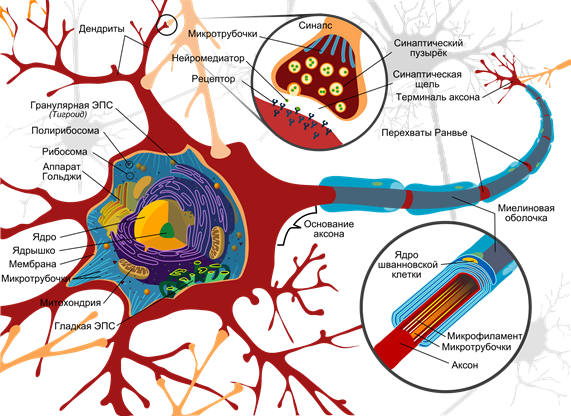
电路神经元(马里亚纳鲁伊斯比利亚雷亚尔) i>的
表面覆盖有一个神经元分支过程 - 树突。其他神经元的轴突邻接枝晶的末端。他们的关节称为突触。通过突触神经元的相互作用可以响应于输入信号,并在某些情况下,以产生其自己的动量,称为尖峰。
信号传输的突触发生过的化学物质称为神经递质。当沿轴突神经冲动进入突触,它会释放神经递质分子囊泡这种突触的特质。上的神经元的膜,该接收信号具有蛋白质分子 - 受体。受体的神经递质进行交互。
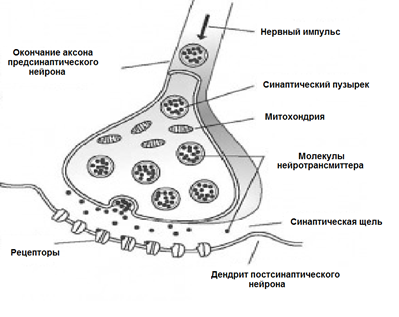
突触 I>
位于突触间隙受体是离子型。它强调的事实,它们是相同的离子通道,能够移动离子。神经递质作用于受体,使他们的离子通道开放。因此,膜被去极化,极化或 - 取决于哪些通道受到影响,并且,相应地,这种类型的突触。在兴奋性突触开放的渠道渗透阳离子进入细胞 - 细胞膜去极化。在抑制性突触开放的渠道,提供负离子,导致细胞膜超极化。
在某些情况下,突触可以改变他们的灵敏度被称为突触可塑性。这导致一个事实,即一个神经元突触成为易感性外部信号之间的不同。
同时,将神经元的突触接收多个信号。抑制性突触拉膜电位向电荷的笼子内的积累。激活突触,相反,试图化解神经元(如下图所示)。
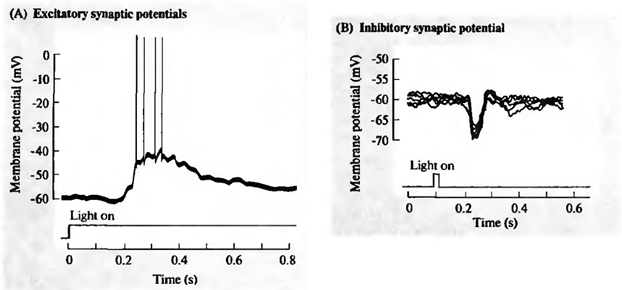
激励(A)和抑制(B)视网膜神经节细胞(J.尼科尔斯。马丁R.华莱士B.,P.福克斯,2003年) i>的
当总活度超过起始的阈值,放电,称为动作电位或尖峰。穗 - 一个神经元,其产生的电脉冲的膜的一个尖锐的去极化。产生动力的整个过程持续约1毫秒。然而,无论是长度也不是脉冲的幅度不依赖于如何进行强烈的理由中高级(见下面的图)。
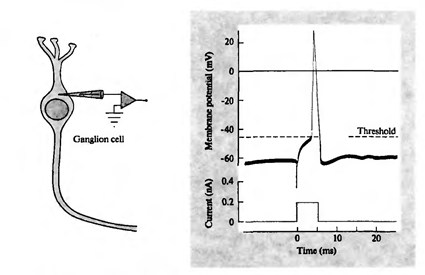
加入动作电位神经节细胞(J.尼科尔斯。马丁R.华莱士B.,P.福克斯,2003年) i>的
斯派克之后离子泵提供神经递质的再摄取和清除突触间隙。在不应期,秒杀后到来,神经元不能产生新的冲动。此期间确定的最大振荡频率,其能够神经元。
出现如在突触活动的结果尖峰,称为诱发。尖峰的频率所引起的重复编码如何以及在输入信号对应于设置的神经元突触的灵敏度。当信号落在敏感突触激活的神经元,并且它不与到达抑制性突触的信号干扰时,神经元的响应达到最大。是受神经元的信号特性中描述的图像被称为刺激。
当然,在神经元的想法是不必要的简单化。某些神经元之间的信息可以被传输不仅尖峰,而且还通过连接通道内的内容,并直接发送的电势。这种延伸被称为渐进的,但化合物本身被称为电突触。树突,根据从所述电池主体的距离被分成近端(接近)和远端(远程)。远端部分可形成枝晶,其充当半元素。除了突触途径有引起代谢性粘连突触外的激励机制。此外也有诱发活动和自发活动。最后,大脑的神经元由神经胶质细胞,这也对正在运行的进程一显著影响包围。
漫长的进化之路创造了自己的工作中使用的大脑许多机制。他们中的一些可以通过本身可以理解,这意味着只考虑相当复杂的相互作用,当其他变得清晰。所以不要拿上面的描述是由神经元作为无遗。去更深的模式,首先要处理的神经元的“基本”的性质。
1952年,艾伦·劳埃德·霍奇金淋巴瘤和安德鲁赫胥黎作了描述,决定在巨型乌贼轴突(霍奇金,1952年)的生成和神经信号的传输电的机制。什么被评为诺贝尔生理学或医学奖于1963年。型号霍奇金 - 赫胥黎介绍常微分方程的神经系统的行为。这些方程对应于活性介质中的自振荡的过程。他们考虑到该组组件,其中每一个具有其在真实的生物物理小区对应物(下图)。离子泵对应于电流源I <子> P 子>。细胞膜的内脂层形成的电容器具有电容C <子>米子>。离子通道突触受体提供电导率克<子>Ñ子>,这取决于输入信号随时间t的变化,以及膜电位V的总价值的膜孔的泄漏电流产生导体克<子>→子> 。离子对离子通道的运动发生电化学梯度的影响下,对应于一个电压源与一个电动势E. <子>Ñ子>和E <子>→子>。
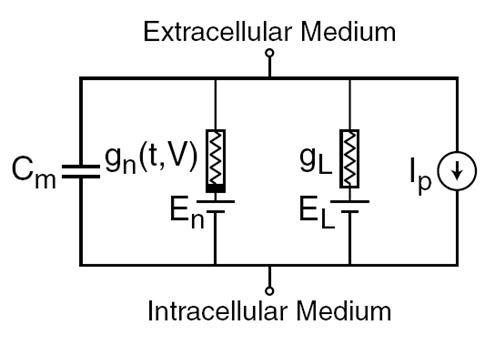
霍奇金淋巴瘤的模型的主要组成部分 - 赫胥黎 I>
当然,与创建神经网络有一个愿望,以简化将神经元的模型,只留下最基本的属性。最有名的和流行的简化模型 - 一个人工神经元麦卡洛克 - 皮茨,在20世纪40年代初开发(J.麦卡洛克,W.皮茨,1956年。)
。
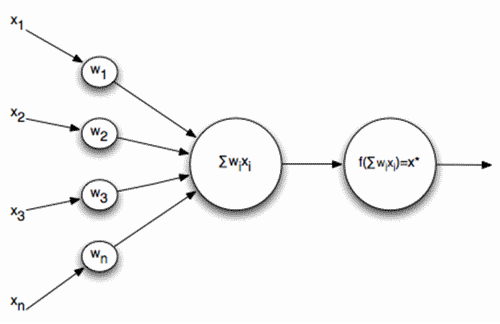
正式的神经元麦卡洛克 - 皮茨 I>
神经元信号的输入是。这些信号被相加均衡。旁边的该线性组合使用一些非线性激活函数,例如,S形。经常被用来作为一个S型的物流功能:
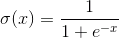
的物流功能 I>
在这种情况下,活动记录为一个正规的神经元
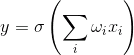
作为这样的神经元的结果变为阈加法器。在神经元输出信号的足够陡阈值函数 - 0或1的输入信号的加权和与神经元的权值 - 两个图像的卷积:所述输入信号的图像,并通过神经元的权重所描述的图像。这些图像的越高,更精确的匹配的卷积的结果。即,神经元实际上是在决定了信号如何类似于所提供的图像,记录在其突触。当卷积的值超过一定的水平和阈值函数被切换到1,它可以被解释为,他了解到呈现所述图像的神经元的一个强有力的声明。
真正的神经细胞的确在某种程度上类似于神经元麦卡洛克 - 皮茨。尖峰的幅度不依赖于什么信号,他们被称为突触。秒杀,无论有或没有。但真正的神经元的刺激不是一个单一的脉冲和脉冲序列作出响应。脉冲的频率,更高的准确知道典型的神经元的图像。这意味着,如果我们建造的加法器这样的阈值是一个神经网络,它是在一个静态的输入信号,并提供在输出端的结果,但结果会远离如何实际的神经元的再生。为了使神经网络对生物原型,我们需要模拟的重放信号的一个给定的时间和频率特性的动态。
但你可以走另一条路。例如,可以选择的神经活动的广义特征,其对应于它的脉冲的频率,即尖峰超过一定的时间周期的数目。如果你去这说明,人们可以想像一个神经元作为一个简单的线性组合。
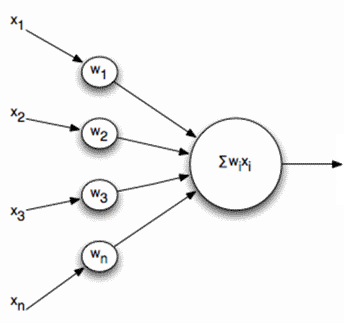
线性组合 i>的
和输出信号,分别针对此类神经元的输入不dihatomichnymi(0或1),并表示在某些标量。时间的激活函数被记录为
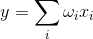
线性加法器不应该被看成是根本不同的与脉冲的神经元相比较,它只是允许建模或描述去的时间间隔更长。虽然脉冲描述更准确,过渡到一个线性组合是合理的在许多情况下模式的强烈简化。此外,一些重要的性能是困难的以脉冲神经显而易见线性组合来辨别。
续
参考
阿列克谢Redozubov (2014)
来源: habrahabr.ru/post/214109/