3355
小事的思想或物品上的树突棘
几个月前发表了一系列的文章,题目是 «思维逻辑»。乐观地计划在一两个月后,继续吧。但生活做出了调整。造型图案波模型壳给了有趣的结果,使我们不得不推迟一切,包括周期写入Habra的延续。
然而,就在不久前,我写了并张贴在预印本文章( http://arxiv.org/abs/1406.6901 )。在某些方面,它可能是有趣的,那些谁以前在波模型感兴趣。回想一下,关键模型 - 一个声明中表示,神经元能够记忆和学习而不是一个单一的形象,描述了他的突触权重,但也有大量的其他的比图像信号。当然,一个神经元的这种并发症是不符合现有的许多理论和需要的不仅仅是一个认真的研究。下面,我,只是,并尝试描述那些支持我的模型的参数给出之一。
这篇文章不应该被视为一个连续的循环,这是相当一个前传吧。至于我自己,我称这种说法对树突棘的关键作用。
我们首先用公知的复发。
在内部和神经元的电势差的外部之间的静止状态 - 约70毫伏的膜电位。它是由蛋白质分子充当离子泵构成。其结果是,该膜呈现的量,负电荷的细胞内积累,而阳性以外的偏振。
表面覆盖有一个神经元分支过程 - 树突。对神经元和树突的主体是邻近其轴突其他神经元的神经末梢。他们的关节称为突触。通过突触神经元的相互作用能响应输入信号,并在某些情况下生成其自己的动量,称为尖峰。
在突触信号传输情况,通过神经递质的释放。当沿轴突神经冲动进入突触前终端,它释放的突触小泡的分子神经递质是典型的这种突触。上,其接收与一种神经递质相互作用所放置的信号受体的一个神经元的膜。
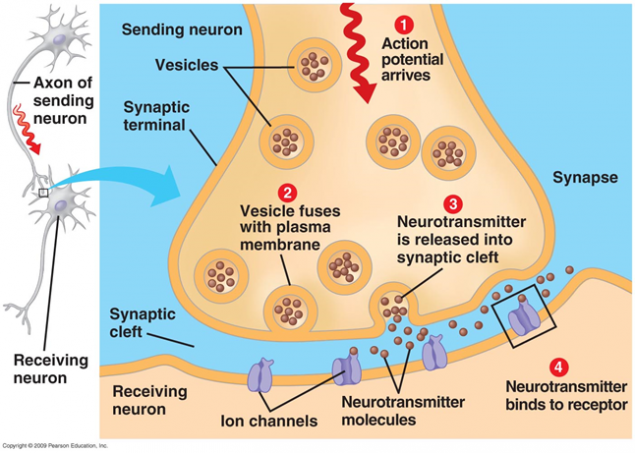
图1.突触
位于突触间隙受体大多是离子型。也就是说,它们是相同的离子通道,通过该神经元的膜移动至离子的能力。神经递质作为受体的离子通道开放。其结果是,膜去极化,或者超极化 - 这取决于通道的影响,并且因此,任何类型的突触。在兴奋性突触打开通道,主要是可渗透阳离子在细胞内 - 膜去极化。在抑制性突触明渠引出细胞,导致细胞膜超极化的阳离子。
在某些情况下,突触可以改变其灵敏度被称为突触可塑性。这导致这样的事实,一些突触变得越来越他人不易受外部信号。
同时,神经元突触收到很多信号。抑制性突触移膜电位向笼子内的电荷累积。激活突触,相反,试图化解神经元。当总数超过起始,放电的门槛去极化,称为动作电位,或尖峰。
神经递质释放后,特别机制确保其回收和再摄取,从而导致突触间隙和周围的突触的空间间隙。在不应期,来秒杀后,神经元是不能产生新的推动力。此周期的持续时间决定了振荡,其能够一神经元的最大频率。
我们现在描述的鲜为人知的事实。
当动作电位传播,沿轴突,神经元到达收件人,它会导致神经递质释放到突触间隙。这些介质确定突触的接收信号的神经元的膜电位的总体变化的贡献。但介质的一部分落入突触间隙外面和填充由周围神经元和神经胶质细胞所形成的空间。这种现象被称为溢出(溢出(英) - 溢出,溢出)(库尔曼,2000)。此外,介质被发射nonsynaptic轴突末梢和神经胶质细胞(图2)。神经递质的突触浓度外面比在突触间隙小得多。然而,它在隐藏利益大众这些“rasteksheysya”的神经递质。
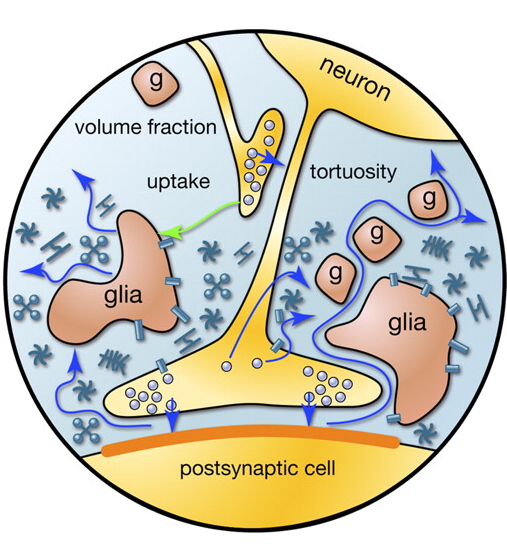
图2.源介质的突触间隙外(Sykova E.,马泽尔吨,Vagrova L.,Vorisek一,Prokopova-Kubinova S.,2000)
让我们试着估计的源发射的突触外神经递质的数量和结构。对于这一点,我们使用皮质的参数的定量估计,如下表所示(表1)(Braitenberg五,Schuz A。,1998)。
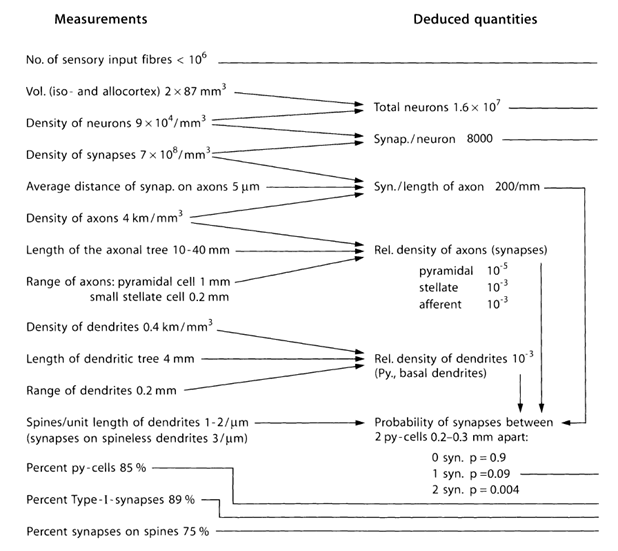
( - 锥体细胞,I型 - PY-细胞锥体细胞之间的突触)在小鼠大脑的研究中获得的参数表1汇总表(Braitenberg五,Schuz A.,1998)
让我提醒你,大部分突触(90-95%)是不是在神经元和树突的身体。树突 - 薄的分支过程形成的神经元的所谓树突树。下面树突树的数字为黑色,灰色和轴突概述。对于不同类型的神经元形成树突的树木都不同,但总的原则是保留:树突树由一组分支流程,具有神经元突触连接的最大密度落在一个小的空间区域。对于基本类型的神经元,它是约200μm(图3,图4)。
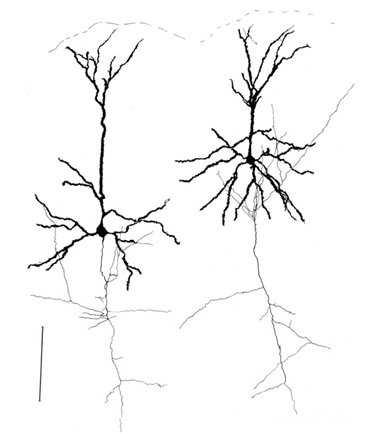
星状神经线图3.结构 - 0.1毫米(Braitenberg,1978)
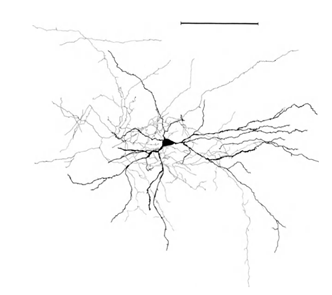
图4.结构锥体神经元,统治者 - 0.1毫米(Braitenberg,1978)
神经元的轴突分枝形成触点(突触)与其它神经元的树突。对树突突触之间的平均距离 - 0.5微米。对轴突突触之间的平均距离 - 5微米,即10倍以上。毫不奇怪,轴突获得约10倍以上的树突。
位于树突突触即锥体细胞(图5)的最有特点的最(75%)。
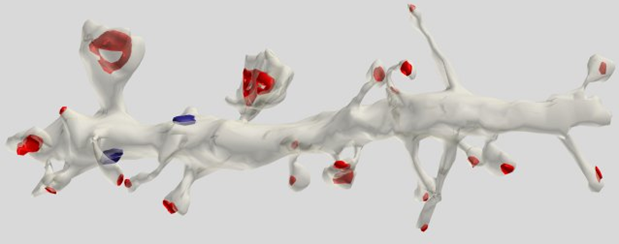
图5.段锥体细胞的枝晶。红色标记的突触上的刺,蓝色 - 树突状干线(博士克里斯汀·M·哈里斯)
真实的解剖和生理数据的基础上进行的计算机模拟表明,例如,谷氨酸盐可能超出突触间隙延伸足以内可比激活NMDA受体与(鲁萨科夫DA相邻的突触(0.5微米)之间的距离的量,库尔曼DM,1998)。它可以假设,溢出后显著浓度递质发生在1-2微米的数量级的枝晶长度的部位。在此网站可以位于属于枝晶约二至四个突触。
如果采取5微米(图6)的枝晶长度的一部分,在其上突触的预期数量将是大约10
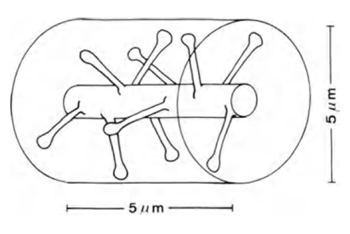
图6.情节枝蔓(Braitenberg五,Schuz A.,1998)
但是神经元的树突一些分支是紧密联系在一起的其他神经元的分支。它们非常接近测试对方。由于属于同一枝蔓树突突触的高度,可以更接近于另一个比他自己的突触的枝晶的表面上。
如果突触被突触的密度分布的7×10 8功能 sup> /毫米 3 SUP>命中均匀地分布于皮质,5mm的高度的圆柱形体积和直径为5毫米,太(上图)的空间至约100突触。即,比直接位于该分支本身的10倍以上。实际上,大脑体积的主要部分由神经胶质细胞和体内,这进一步增加神经突触的填充密度的神经元所占据。然而,神经元采用不同的神经递质,其也应该被考虑。
现在,试着去理解这种结构关系的含义,密度分布突触外神经递质的条款。对于这一点,我们使用一个简化模型。采取有条件的周围神经元的数量,并列举其成员的神经元。每个神经元将是如下:
•与选定的神经元树突多突触联系;
•“适合”,也就是,在那里它与其它神经元位于所选择的神经元枝蔓附近突触地方的数目。
想象一下,树突树的长枝与均匀分布的常规来源(图7)。有关此主题的每个源,您可以指定从周围空间的神经元的数目,对它负责。每个环境的神经元将具有源的多个联系人,沿着树突随机分布的。我们代表的元素ð I SUB>此相关向量D。
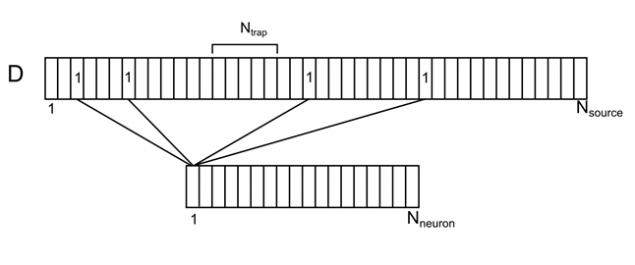
图7的相关性周围神经元的和其上的枝晶
接触
我们引入符号:
ñ<分>神经元 SUB> - 数量周边
神经元 ñ<分>源 SUB> - 源数以单个神经元
ñ<分>陷阱 SUB> - 多种渠道,创造神经递质的浓度水平(突触陷阱)
现在假设环境的某些特定神经元秒杀。这可以被视为一个信号,该信号可用来观看我们的神经元。表示n <分> SIG SUB> - 激活神经元的数量,创造了一个信息信号。我们写这个信号的二进制向量S.
对枝晶,但在最极端的所有位置,我们假定介体的密度由式
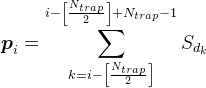
例如,对于在下面图中所示的信号,突触密度在标记的陷阱是2(来自第一和神经元的第四信号的总和)。
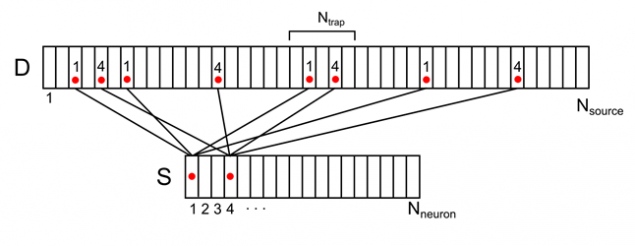
图8显示两个神经元的环境对树突树(只有部分债券和编号)活动
对于任何任意的信号可以计算出介质上的枝晶的图案的密度。这个密度范围从0到N 陷阱 SUB>。最大值将会实现,如果所有的来源是活跃,形成突触陷阱。
我们使用的参数实大鼠皮层(Braitenberg五,Schuz A.,1998)的特征的平均值。在此基础上,我们可以得到下面的模型参数:
ñ<分>神经元 SUB> = 650
ñ<分>源 SUB> = 25000
ñ<分>陷阱 SUB> = 15
我们假设该信号是由活动的编码,例如,皮质的神经元的1.5%,而
ñ<分> SIG SUB> = 10
这是很容易计算的概率选自N <子>的任意信号sig 分>单元,存在于树枝状晶体,其中所述介体的密度会完全K.对于一个给定的概率参数中的至少一个位置具有以下的值(表2):
<表> K TD> P TD> TR> 0 TD> 0,984 TD> TR > 1 TD> 1 TD> TR> 2 TD> 1 TD> TR> 3 TD> 0,996 TD> TR> 4 TD> 0,287 TD> TR> 5 TD> 0,016 TD> TR> 6 TD> 0,001 TD> TR> 7 TD> 0 TD> TR> 8 TD> 0 TD> TR> 9 TD> 0 TD> TR> 10 TD> 0 TD> TR> ... TD> TR> TABLE>
表2中。表查找至少一个凝汽阀具有给定密度的概率。第一列 - 在陷阱活动源所需要的数量。第二 - 找到至少一个枝晶位置的概率在那里将是数字的活性来源
也就是说,对于是接近皮层的实际配置的参数的这些值,对于任何环绕信号,影响约1.5%的神经元,以下方法:
•发现约1.6%的神经元,其上存在的陷阱,其中所述信号越过轴突的50%的树突;
•几乎每个神经元有陷阱,相交的信号轴突中的至少30%。
这一结果的意义是非常有趣的。假设在皮层中的信息以某种方式编码的一个相对小的数目(N <子>签名分>)同步活性紧凑地布置的神经元。我们不是在谈论大脑的整体活动,并在小体积中的信息流程,在神经元的编号从1到美国ñ<分>神经元 SUB>的。假定的码字S中的数目是有界的,并形成特定的词汇量T N <子>字典子>。就可以计算出相同的地方“回应”立即分成两个信号的概率。这个计算的下表万信号的词典的结果。
<表> K TD> P <分>错误 SUB> TD> TR> 3 TD> 0, 00 399 TD> TR> 4 TD> 1,05E-05 TD> TR> 5 TD> 1,89E-08 TD> TR> 6 TD> 2,33E-11 TD> TR> 7 TD > 0 TD> TR> ... TD> TR> TABLE>
的独特性突触陷阱,在不同层次的神经递质浓度
表3.表中断概率
事实证明,如果K = 3的陷阱有一定的选择性,但对错误不能保证,但K = 5,就开始充分对应唯一确定活动的空间格局。让我提醒你,这是不正确的,从一组无限任意信号,而当我们有一组,虽然活动的足够大的离散允许状态。
也就是说,它证明,固有的大脑轴突和树突树的结构的意义是建立在凝汽阀的每个神经元的富空间的树突状表面,即,对应于神经递质的源的所有可能的组合的位置。根据在陷阱中神经递质的浓度,可以判断对组成的一个数附近的神经元的同步活动的信号的空间分布。
而作为位于所谓的代谢型受体神经元的表面,这甚至可以在神经递质的浓度低导致单个神经元的尖峰上,事实证明,这些尖峰,这是所谓的自主神经元可以到数量庞大的不同信号比对了其他回应可以调节神经元的突触。
其实,我们现在可以解释很树突棘与文章开头的目的。这个想法是,所有的突触可直接在枝晶或细胞体放置。这样就不会影响突触的直接工作和他们的能力,有助于诱导活动的发生。但是现在我们可以假设,树突棘的任命 - 是创造空间结构中,不同神经元的突触是“混合”,以便其获得的溢出影响,不仅在其表面的能力,而且周围的邻居树突。这是这里的“委托琐事»。
在大多数的原创文章展示了如何这一切来自于实际的“波的故事,”但是这是一个不同的故事。
并在小增益的末端。原创文章在原俄罗斯。有<一href="https://docs.google.com/document/d/1qStecbZI6hyt_TqN4gdUegSNhYJ1g83R9sJr7t7Fq4g/edit#heading=h.vk35vajr6e2l">перевод英语,取得梅德Shabanov (为此他非常尊重),但他(翻译)是远远不够完善。如果任何人有机会看到并指出了错误,会非常感激。所引用的文件是开放给所有的评论。
阿列克谢Redozubov
引用文章原始
1.(2014年)。从人类连接组项目获得: www.humanconnectomeproject.org
2.(2014年)。从ALLEN小鼠脑连接图谱检索: connectivity.brain-map.org/
3.布卢姆,B. H.(1970)。空间/时间权衡的哈希允许的误差进行编码。在ACM的T. 13通信(7),422-426。
4. Braitenberg五,Schuz A.(1998)。皮质:统计和神经元的连接,第二版的几何形状。
5. Braitenberg,V.(1978)。皮质建筑学:一般和面。在M.A.B.火盆和H. Petsche(EDS),大脑皮层建筑学(页443-465)。纽约:Raven出版
。 6.科斯特,H.(1975)。机电压力和pH值对膜结构的影响。生物化学和生物物理学报13; 382(2),142-146
7.博士克里斯汀·M·哈里斯。 (PPB)。突触网络。从 synapses.clm.utexas.edu/ 检索。
8.福岛,K.(1980)。 Neocognitron模式识别不受偏移位置的机制,自组织神经网络模型。生物控制论,36(4),193-202。
9.格罗斯伯格,S.(1987)。竞争学习:从交互激活自适应共振。认知科学N11,23-63。
10.赫布,D.(1949)。行为的组织。纽约:约翰·威利父子
11.何杰金氏,A的。 (1952)。膜电流的定量描述及其应用导和激发神经... J.队医升。 117,500-544。
12. Izhikevich,E.M。(2007)。动力系统在神经科学:兴奋性和爆破的几何形状。伦敦:麻省理工学院出版社
。 13.库尔曼,D。M。(2000)。溢出与谷氨酸和GABA在哺乳动物大脑突触介导的相声。 PROG脑研究,125,339-351。
14.仓本,Y。(1984)。化学振荡,波浪和湍流。多佛出版物。
15. Liibke J.,马克拉姆H。,Frotscher M.,索克曼B。(1996)。次数由层建立5锥体神经元发育大鼠大脑皮层autapses的树突状分布bution:具有相同类的相邻神经元的突触神经支配的比较。神经科学16(10),3209-3218。
16. Malenka R.C.,尼科尔R.A. (1999年)。长时程增强 - 十年的进步?科学285(5435),1870年至1874年。
17.迈克尔·利珀特,Kentaroh高垣,李玮锋徐,黄晓莺,建扬武。 (2007)。方法大鼠皮层活动具有电压敏感染料成像。