3365
Trivia thinking or article on dendritic spines
A few months ago published a series of articles under the title «The logic of thinking» . Optimistically planned to continue it after a month or two. But life has made adjustments. Modeling pattern wave model crust gave interesting results so that we had to postpone everything else, including the continuation of the cycle for writing Habra.
However, not so long ago, I wrote and posted on the preprint article ( http://arxiv.org/abs/1406.6901 ). In some ways it may be interesting to those who previously interested in the wave model. Recall that the key point model - a statement that the neurons are able to memorize and learn not one single image, described his weights of synapses, but also a huge amount of other than that image signals. Of course, such a complication of a neuron is at odds with many of the existing theories and requires more than a serious study. Below, I, just, and try to describe one of those given in the arguments in favor of my model.
This article should not be taken as a continuation of the cycle, it is rather a prequel to it. For myself, I have called this argument about the key role of dendritic spines.
We begin with the well-known recurrence.
In the quiescent state between the inside and the outside of the neuron potential difference - the membrane potential of around 70 mV. It is formed by the protein molecules that act as ion pumps. As a result, the membrane assumes the polarization for which the negative charge is accumulated within the cells, and positive outside.
The surface is covered with a neuron branching processes - dendrites. To the body of the neuron and the dendrites are adjacent to its axonal endings of other neurons. Their joints are called synapses. Through the interaction of synaptic neuron can respond to incoming signals and under certain circumstances to generate its own momentum, called spike.
Signal transmission at the synapses happens through neurotransmitter release. When a nerve impulse along the axon enters the presynaptic terminal, it releases from the synaptic vesicle molecule neurotransmitters that are typical of this synapse. On the membrane of a neuron that receives the signal placed receptors that interact with a neurotransmitter.
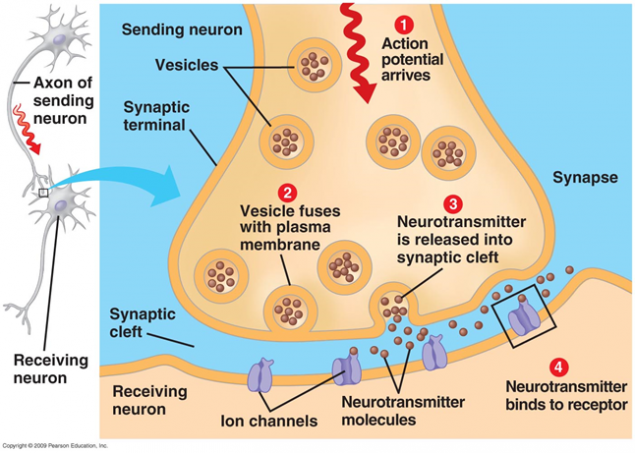
Figure 1. Chemical synapse
Receptors located in the synaptic cleft are mostly ionotropic. That is, they are the same ion channels, the ability to move ions through the membrane of the neuron. Neurotransmitters act as receptors that their ion channels open. As a result, a membrane is depolarized, or hyperpolarizing - depending on which channels are affected, and accordingly, any type of synapse. In excitatory synapses opens channels, mostly permeable cations inside the cell - membrane is depolarized. In inhibitory synapses open channels leading out cations of cells, which leads to membrane hyperpolarization.
In certain circumstances, the synapses can change their sensitivity is called synaptic plasticity. This leads to the fact that some of the synapses become more and others less susceptible to external signals.
Simultaneously, the neuron synapses receive many signals. Inhibitory synapses shift the membrane potential toward the charge accumulation inside the cage. Activating the synapses, on the contrary, try to defuse the neuron. When the total exceeds the threshold depolarization of initiation, a discharge, called an action potential, or spike.
After release of neurotransmitters special mechanisms ensure their recycling and re-uptake, which leads to the clearance of synaptic gap and the space surrounding the synapse. During the refractory period, comes after a spike, the neuron is not able to generate new impulses. The duration of this period determines the maximum frequency of oscillation, which is capable of a neuron.
We now describe the lesser-known facts.
When an action potential propagates along the axon, the neuron reaches the recipient, it causes the release of neurotransmitters into the synaptic cleft. These mediators determine the contribution of the synapse in the overall change in the membrane potential of the neuron that receives the signal. But part of mediators falls outside the synaptic gap and filling the space formed by the surrounding neurons and glial cells. This phenomenon is called spillover (spillover (Eng.) - Overflow, overflow) (Kullmann, 2000). In addition, mediators are emitted nonsynaptic axon terminals and glial cells (Figure 2). The concentration of neurotransmitter synaptic outside is much smaller than in synaptic clefts. However, it is in these "rasteksheysya" neurotransmitters hidden mass of interest.
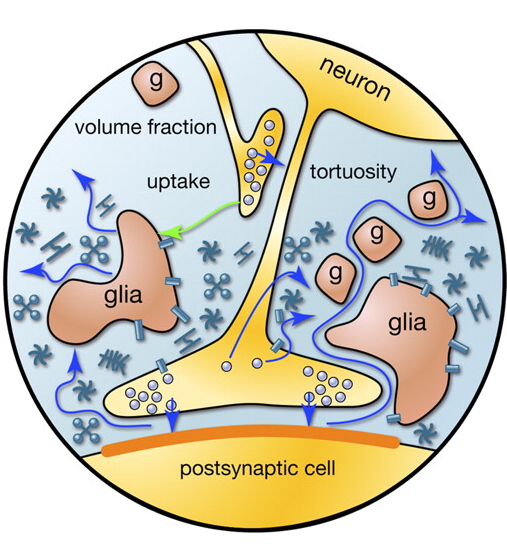
Figure 2. Sources of mediators outside the synaptic cleft (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
Let us try to estimate the number and structure of sources emitting neurotransmitters outside of synapses. For this we use quantitative estimates of the parameters of the cortex, in the table below (Table 1) (Braitenberg V., Schuz A., 1998).
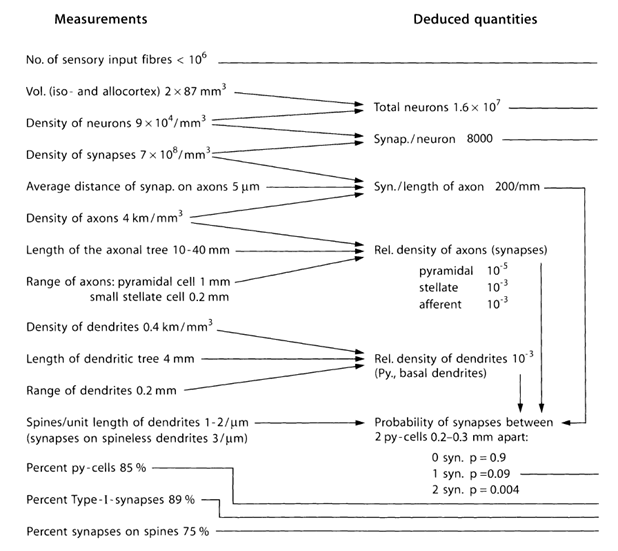
Table 1. Summary table of the parameters obtained in the study of mouse brain (py-cell - pyramidal cell, Type-I - synapses between pyramidal cells) (Braitenberg V., Schuz A., 1998)
Let me remind you that most of the synapses (90-95%) is not on the body of the neuron and its dendrites. Dendrites - a thin branching processes forming the so-called dendritic tree of the neuron. The figures below dendritic trees outlined in black, gray and axons. For different types of neurons form dendritic trees are different, but the general principle is preserved: dendritic tree consists of a set of branching processes, with the greatest density of synaptic connections of a neuron falls on a small spatial region. For basic types of neurons, it is about 200 m (Figure 3, Figure 4).
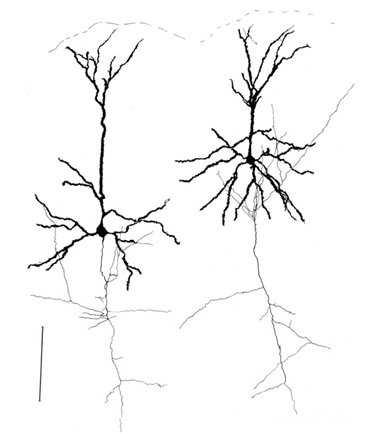
Figure 3. Structure of the stellate neuron line - 0.1 mm (Braitenberg, 1978)
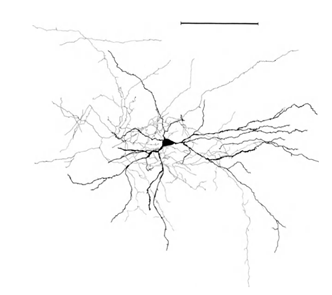
Figure 4. Structure of pyramidal neurons, ruler - 0.1 mm (Braitenberg, 1978)
Branching axons of neurons form contacts (synapses) with the dendrites of other neurons. The average distance between synapses on the dendrites - 0.5 micrometer. The average distance between synapses on axons - 5 micrometers, that is 10 times greater. Not surprisingly, the axons are obtained approximately 10 times longer than the dendrites.
Most (75%) of synapses located on dendritic spines that is most characteristic of pyramidal cells (Figure 5).
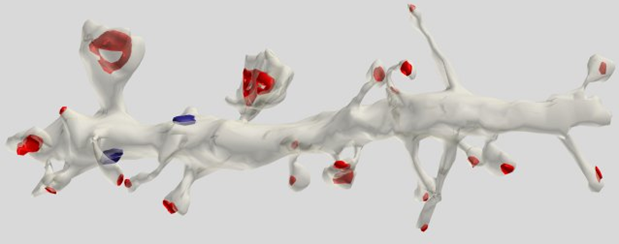
Figure 5. Segment dendrite of pyramidal cells. Red marked synapses on spines, blue - on dendritic trunk (Dr. Kristen M. Harris)
Computer simulations performed on the basis of real anatomical and physiological data showed that, for example, glutamate may extend beyond the synaptic cleft in amounts sufficient to activate NMDA receptors within comparable with the distance between neighboring synapses (0.5 microns) (Rusakov DA, Kullmann DM, 1998). It can be assumed that a significant concentration of neurotransmitters after spillover occurs at the site of the dendrite length of the order of 1-2 microns. On this site can be located about two to four synapses belonging to the dendrite.
If you take a portion of the dendrite length of 5 m (Figure 6), the expected number of synapses on it will be about 10.
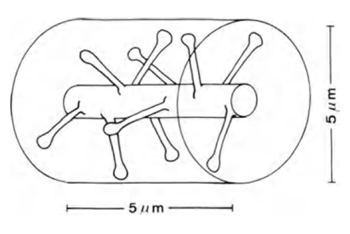
Figure 6. Plot of dendrite (Braitenberg V., Schuz A., 1998)
But some dendritic branches of neurons is closely intertwined with the branches of other neurons. They tested each other in very close proximity. Due to the height of dendritic spines synapses belonging to the same dendrite, may be closer to the surface of the dendrite of another than his own synapses.
If synapses were uniformly distributed in the space of the cortex, the cylindrical volume of height of 5 mm and a diameter of 5 mm, too (picture above) at a density distribution of synapses 7x10 8 sup> / mm 3 sup> hit to about 100 synapses. That is, 10 times more than what is directly located on the branch itself. In fact, a substantial part of the brain volume occupied by glial cells and neurons of the body, which further increases the packing density of synapses. However, neurons employ different neurotransmitters, that too should be considered.
Now try to understand the meaning of such a structure relationships in terms of the density distribution extrasynaptic neurotransmitter. For this we use a simplified model. Take conditional surrounding neurons volume and enumerate its member neurons. Each of these neurons would be:
• multiple synaptic contacts with the selected neuron dendrite;
• number of "fit" that is, places where it synapses with other neurons are located in the vicinity of the selected neuron dendrite.
Imagine dendritic tree as a long branch with uniformly distributed conventional sources (Figure 7). For each source on this thread, you can specify the number of neurons from the surrounding space, is responsible for it. Each of the neurons of the environment will have multiple contacts of source, randomly distributed along the dendrites. We denote this correlation vector D with elements d i sub>.
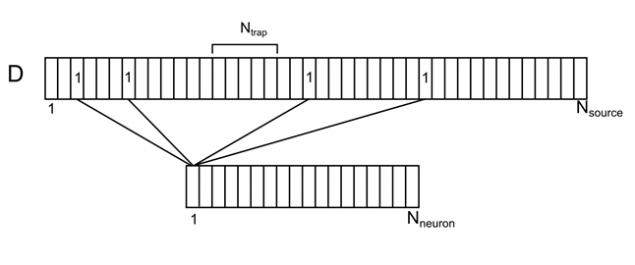
Figure 7. Correlation of the surrounding neurons and their contacts on dendrite
We introduce the notation:
N neuron sub> - number of neurons surrounding
N source sub> - number of sources to a single neuron
N trap sub> - number of sources, creating a density level of neurotransmitters (synaptic trap)
Now suppose that some neurons of the environment given spike. This can be taken as a signal that is available to watch our neuron. Denote N sig sub> - the number of active neurons, creating an information signal. We write this signal binary vector S.
For all positions on the dendrite but the most extreme, we assume the density of the mediator by the formula
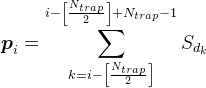
For example, for the signal shown in the figure below, the synaptic density in the marked trap is 2 (the sum of the signals from the 1st and 4th of neurons).
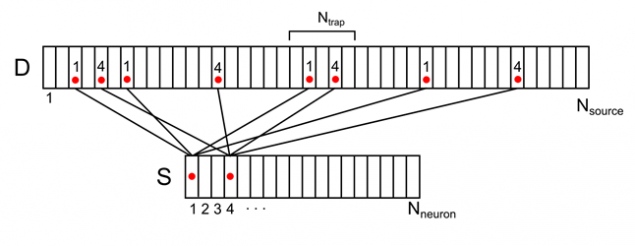
Figure 8. Display of the activity of two neuronal environment on dendritic tree (only part of the bonds and numbering)
For any arbitrary signal can calculate the density of the pattern of mediators on the dendrite. This density will range from 0 to N trap sub>. The maximum value will be achieved if all the sources were active, forming synaptic trap.
We use the average values of parameters characteristic of the real rat cortex (Braitenberg V., Schuz A., 1998). Based on these, we obtain the following model parameters:
N neuron sub> = 650
N source sub> = 25000
N trap sub> = 15
We assume that the signal is encoded by the activity, for example, 1.5% of the neurons of the cortex, whereas
N sig sub> = 10
It is easy to calculate the probability that an arbitrary signal consisting of N sig sub> units, there is at least one place on the dendrite, where the density of the mediator will be exactly K. For a given probability parameters have the following values (Table 2):
| K | P | ||||||||||||
| 0 | 0, 984 tr > | ||||||||||||
| 1 | 1 | ||||||||||||
| 2 | 1 | ||||||||||||
| 3 | 0, 996 | ||||||||||||
| 4 | 0, 287 | ||||||||||||
| 5 | 0, 016 | ||||||||||||
| 6 | 0, 001 | ||||||||||||
| 7 | 0 | ||||||||||||
| 8 | 0 | ||||||||||||
| 9 | 0 | ||||||||||||
| 10 | 0 | ||||||||||||
| ... Table 2. Table of the probability of finding at least one trap with a given density. First column - the required number of active sources in the trap. Second - the probability of finding at least one dendrite place where it will be a number of active sources That is, for those values of the parameters that are close to the actual configuration of the cortex, for any surround signal, affects about 1.5% of the neurons, the following: • find about 1.6% of the neurons, which exists on the dendrites of the trap, where the signal crosses 50% of axons; • Almost every neuron there are traps, which intersects at least 30% of the signal axons. The meaning of this result is very interesting. Assume that the information in the cortex is somehow encoded synchronous activity of a relatively small number (N sig sub>) is compactly arranged neurons. We are not talking about the whole activity of the brain, and of the information processes in a small volume, where the neurons are numbered from 1 to us N neuron sub>. Assume that the number of codewords S is bounded and forms a specific vocabulary capacity T N dict sub>. You can calculate the probability that the same place "respond" immediately into two signals. The results of this calculation for a dictionary of 10,000 signals in the table below.
|