3364
Малі думки або статті про дендротичні хребти
Кілька місяців тому під загальним титулом « Логіка мислення». Оптимально планується продовжити його в місяць або два. Але життя зробив свої налаштування. Моделювання шаблонно-хвильової моделі кори давали так цікаві результати, які нам довелося перенести все ще на деякий час, в тому числі написання продовження циклу для концентратора.
Не довге тому я писав і додрукував статтю (http://arxiv.org/abs/1406.6901). У деяких випадках може бути цікавим для тих, хто раніше зацікавився моделлю хвилі. Нагадуємо, що ключова точка моделі є заявою, що нейрони здатні запам'ятати і розпізнати не одне зображення, описане вагами його сипс, але і величезною кількістю інших сигналів, які відрізняються від цього образу. Звісно, це ускладнення нейрона на коефіцієнтах з багатьма існуючими теоріями і вимагає більш серйозного обґрунтування. Далі я спробую описати один з аргументів на користь моєї моделі.
Ця стаття не повинна бути прийнята як продовження циклу, але досить притискати до неї. Для себе я зателефоную цю причину про ключову роль дендротичних хребтів.
Почнемося повторювати добре відомі.
В іншому випадку є потенційна відмінність між внутрішнім і зовнішнім середовищем нейрона – мембранний потенціал близько 70 мілівольтів. Утворюється молекули білків, які працюють як іонні насоси. В результаті мембрана набуває поляризацію, в якій негативний заряд накопичується всередині клітини і позитивний заряд зовні.
Поверхня нейрона покрита розгалуженими процесами - дендритами. Нейроне тіло і його дендрити прилягають до осінніх кінцівок інших нейронів. Місця їх з'єднань називають сипсами. Через сиптичну взаємодію нейрон здатний реагувати на вхідні сигнали і, за певних обставин, генерувати власний імпульс, який називається спрейкою.
Зняття нейротрансмітаторів відбувається при передачі сигналу. При аксоновому нервовому імпульсі надходить до пресинаптичного терміналу, він випускає молекули нейротрансміттера, характерні для сипсу від сиптичних вушок. На мембрані нейрона отримують сигнал, є рецептори, які взаємодіють з нейротрансмітаторами.
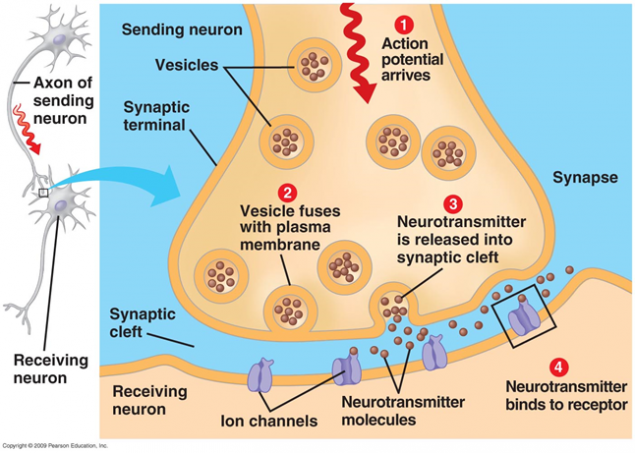
Малкa 1. Хімічний снайпер
Рецептори, розташовані в сиптиці, в основному ionotropic. Це також іонні канали, які можуть пересуватися іонів через мембрану нейрона. Нейротрансмітери впливають на рецептори, щоб їх іонні канали відкриті. В результаті мембрана або деполяризація або гіперполяризація, в залежності від яких вражаються канали і, відповідно, який тип засмаги це. У збудливих знімках з'являються канали, в основному пропускаються змії в клітинку - мембрана деполяризована. У гальмівних знімках відкриваються канали, які знімають з целюнки, що призводить до гіперполяризації мембрани.
При певних обставинах можна змінити чутливість, яка називається синоптичним пластичністю. Це викликає деякі знімки, щоб стати більшими та іншими, щоб стати менш схильними до зовнішніх сигналів.
У той же час багато сигналів наділяються на знімки нейрона. Смола гальмування зсуву потенціалу мембрани в напрямку накопичення заряду всередині клітини. Активувати сніпки, навпаки, спробувати випускати нейрон. При загальній деполяризації перевищить поріг ініціації, відбувається виділення, що називається потенціалом дії або походом.
Після виходу нейротрансмітерів, спеціальні механізми забезпечують їх використання і перезапуск, що призводить до очищення спинного клаптяви і навколишнього простору. Під час вогнетривкого періоду слідуючи щуки, нейрон не здатний генерувати нові імпульси. Тривалість даного періоду визначає максимальну частоту генерації, яка здатна нейрон.
Давайте описати менш відомі факти.
Коли потенціал дії, пропагуючи по осі, досягає нейрона одержувача, це викликає виділення нейротрансмітаторів в симпатичну клітку. Ці медіатори визначають внесок синапсу до загальної зміни мембранного потенціалу нейрона, що отримує сигнал. Але деякі медіатори падають за межі спинного клацання і поширення через простір, утворений нейронами і навколишніми гліальними клітинами. Це явище називається розпіловер (переповнення) (Кульман, 2000). Крім того, медіатори випромінюють несинаптичні аксонні термінали та гліальні клітини (рис. 2). Концентрація нейротрансмітаторів за межами снайперів значно менше, ніж у сиптичних клацках. Тим не менш, це в цих нейротрансмітерах, які багато цікавих речей лука.
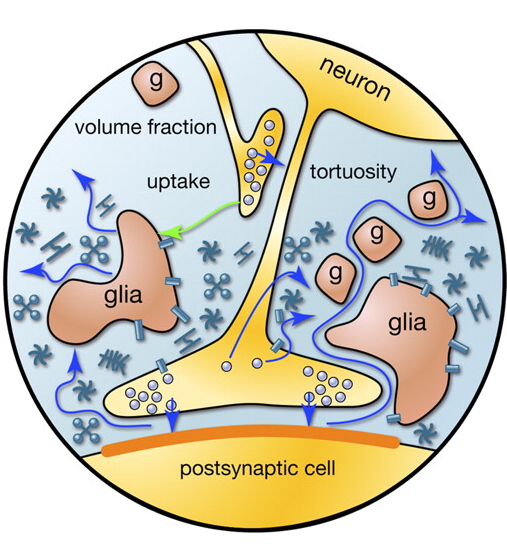
Малкa 2. Джерела медіаторів за межами симпатичного cleft (Сикова Є., Мазель Т., Вагрова Л., Ворісек І., Прокопова-Кубінова С., 2000)
Давайте спробуємо оцінити кількість і структуру джерел, які випускають нейротрансмітери за межі снапів. Для цього скористайтеся кількісними оцінками кортичних параметрів, наведених в таблиці нижче (Таблиця 1) (Браїтенберг В., Шуз А., 1998).
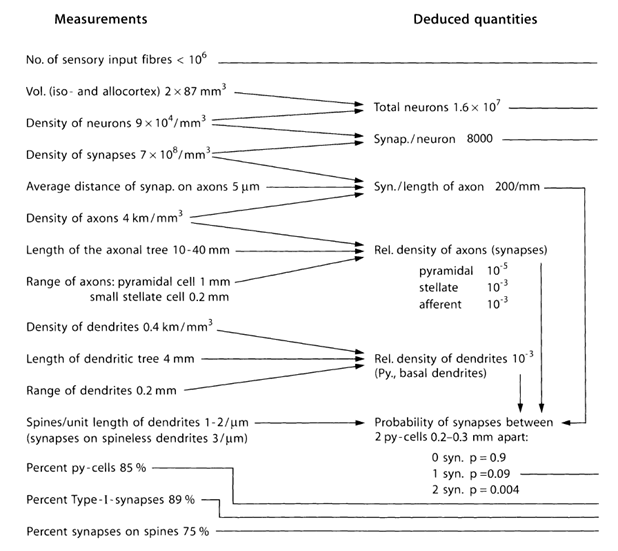
Таблиця 1. Резюме таблицю параметрів, отриманих шляхом вивчення головного мозку миша (py-cell - пірамідальна клітина, Type-I - synapses між двома пірамідальними клітинами) (Брайтенберг В., Шуз А., 1998)
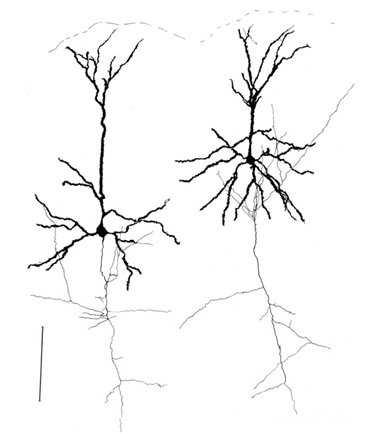
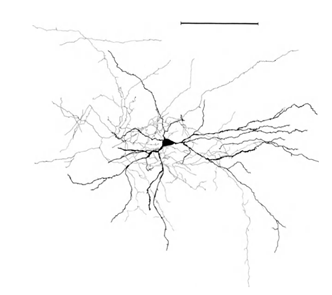
Нагадуємо, що більшість сипсів (90-95%) не є тілом нейрона, але його дендрити. Dendrites - це тонкі розгалужені процеси, які утворюють так звану нейронну дендротичну дерево. На рисах нижче, дендротичні дерева виділяються в чорному і осьовому кольорі сірий. Для нейронів різних типів різні форми дендрітичних дерев, але загальний принцип залишається: дендрітична дерево складається з багатьох розгалужених процесів, з найбільшою щільністю сиптичних з'єднань нейрона, що відбуваються в невеликій просторій зоні. Для основних типів нейронів це близько 200 мкм (рис. 3, рис. 4).
Малкa 3. Структура столярного нейрона, лінійка – 0,1 мм (Браїтенберг, 1978)
Малкa 4. Структура пірамідальної нейрона, лінійка – 0,1 мм (Брайтенберг, 1978)
У відділенні аксони нейронів утворюють контакти (синопси) з дендритами інших нейронів. Середня відстань між сипсами на дендритах становить 0,5 мікрометри. Середня відстань між снайперами на ось 5 мікрометрів, які в 10 разів більше. Не дивно, що ось близько 10 разів довше, ніж дендрити.
Більшість (75%) сипсів розташовуються на дендротичних хребтах, які найбільш характерні для пірамідальних клітин (рис. 5).
447083 р.
Рисунок 5. Відрізок пірамідних клітин. Червоні шоу симпатики на хребтах, сині на дендрицькому стовбурі (Dr. Kristen M. Harris)
Комп'ютерні імітації на основі реальних анатомічних і фізіологічних даних показали, що, наприклад, глутамат може поширюватися за межі симпатичного клітка в кількості, достатній для активації рецепторів НМДА в радіусі, що порівняно з дистанцією між сусідніми сипсами (0.5 мкм) (Rusakov D. A., Kullmann D. M., 1998). Можна припустити, що значна концентрація нейротрансмітаторів після шпловеру спостерігається на місці дендриту довжиною близько 1-2 мкм. На такому сайті можна розмістити близько двох або чотирьох знімків, що відносяться до цього дендрайту.
Якщо ви берете дендрайт 5 мкм довга (рис. 6), очікувана кількість сипсів на ньому буде близько 10.
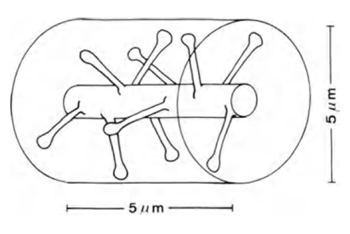
Малкa 6. Dendrite сайт (Брайтенберг В., Шуз А., 1998)
Але дендрітичні гілки деяких нейронів тісно переплітаються гілками інших нейронів. Поблизу в безпосередній близькості. У зв'язку з висотою дендротичних хребтів, знімки, що відносяться до одного дендриту, можуть бути ближче до поверхні іншого дендриту, ніж власних сипсів.
Якщо сипси були рівномірно розподілені в космосі, то близько 100 сипсів впадуть в циліндричний об'єм з висотою 5 мкм і діаметром також 5 мкм (рис вище) з сипучою щільністю 7x108/мм3. Це 10 разів більше, ніж що знаходиться безпосередньо на гілці. По суті, значна частина об'єму мозку зайнята гліальними клітинами і нейронними тілами, що додатково збільшує щільність пакету сипсів. Однак нейрони працюють з різними нейротрансмітаторами, які також слід враховувати.
Тепер ми спробуємо зрозуміти значення такої структури з'єднань з точки зору розподілу густини екстрасинаптичного посередника. Для цього ми використовуємо спрощену модель. Прийміть обумовлений об'єм, що оточує нейрон і занурює нейрони. Кожен з цих нейронів буде мати:
кілька синоптичних контактів з дендритом обраного нейрона;
кілька "приховує", тобто місця, де його знімки з іншими нейронами будуть розташовуватися в безпосередній близькості від дендриту обраного нейрона.
Уявіть дендротичну дерево як одне довге відділення з рівномірно розподіленими звичайними джерелами (рис. 7). Для кожного джерела на цій галузі можна вказати кількість нейронів з навколишнього простору, відповідального за неї. Кожен з нейронів в середовищі буде мати кілька вихідних контактів випадковим чином розподілених по дендриту. Визначаємо це відношення до вектора D з елементами ді.
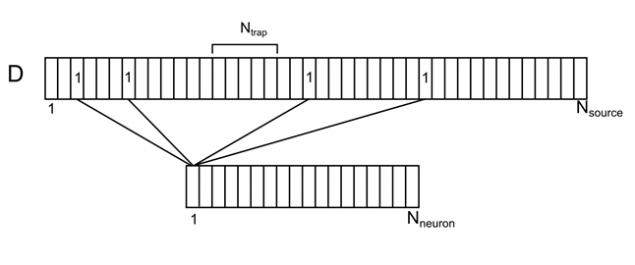
Малкa 7. Корреляція оточуючих нейронів та їх контактів на дендритах
Давайте знайомимо етикетки:
Не нейрон – кількість нейронів у середовищі
N джерело – кількість джерел для одного нейрона
Н пастка - кількість джерел, які створюють рівень щільності нейротрансмітаторів (синаптична пастка)
Тепер припустимо, що кілька нейронів в поході навколишнього середовища. Це може сприйматися як сигнал, доступний для спостереження за нашими нейронами. N Sig - це кількість активних нейронів, які виробляють інформаційний сигнал. Напишіть цей сигнал з бінарним вектором S.
Для всіх позицій на дендриті, крім найбільш екстремальних, ми розглянемо щільність медіатора відповідно до формули
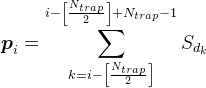
Наприклад, для сигналу, показаного на малюнку нижче, щільність в позначеному сонному пасі буде 2 ( сума сигналів від 1-го і 4-го нейронів).
до 1 2 3 4+
Малкa 8. Показано активність двох середовищних нейронів на дендрицькому дереві (тільки частина з'єднань і нумерації)
Для будь-якого довільного сигналу можна розрахувати щільність медіаторів на дендритах. Ця щільність буде діапазон від 0 до N пастка. Максимальне значення буде досягнуто, якщо активні всі джерела, що утворюють сиптичну пастку.
Ми використовуємо середні значення параметрів, характерних для реальної кори щурів (Браїтенберг В., Шуз А., 1998). На основі них ми отримаємо такі параметри моделі:
N нейрон = 650
N джерело = 25,000
N пастка = 15
Припустимо, що сигнал закодовано активністю, наприклад, 1.5% нейронів у корі, потім
N сіг = 10
Легко розрахувати ймовірність, що для довільного сигналу, що складається з вузлів N, є принаймні одне місце на дендриті, де щільність медіатора точно K. Для заданих параметрів ймовірність приймає такі значення (Таббі 2):
К П 0 0 0 0 0 0 0 0 0
Настільний 2. Імовірність таблиці знаходження принаймні однієї пастки з даної щільності. Перший стовпчик є необхідною кількістю активних джерел в пастку. Друга ймовірність пошуку принаймні одного місця на дендріті, де буде така кількість активних джерел.
Тобто, при значеннях, які близькі до конфігурації реальної кори, для будь-якого об’ємного сигналу, що впливає приблизно на 1,5% нейронів, наступне:
• Є близько 1,6% нейронів, які мають пастку на дендритах, де 50% сигналів перетинаються.
• Практично кожен нейрон має пастки, в яких не менше 30% сигнальних осей.
Значення цього результату дуже цікавим. Припустимо, що інформація в корі якось зашифрована синхронною активністю порівняно невеликого числа (N sig) компактно розташованих нейронів. Ми не говоримо про всі мозкові дії, але про інформаційні процеси в невеликому обсязі, де нейрони нумеровані від 1 до нейрона. Припустимо, що кількість комбінацій коду S обмежена і формує певний словниковий запас Т з ємністю N дикт. Ви можете розрахувати ймовірність того, що одне місце буде реагувати на два сигнали. Результати цього розрахунку на 10 000 знаків у таблиці нижче.
К П помилка 3 0.00399 4 1.05E-05 5 1.89E-08 6 2.33E-11 7 0
Таблиця 3. Таблиця ймовірності порушення унікальності сиптичних пасток на різних рівнях щільність медіатора
Виходить, що на K = 3 пастки мають певну вибірковість, хоча вони не гарантують від помилки, але вже на K = 5 вони починають відповідати певному просторовому шаблону діяльності. Нагадуємо, що це правда не для довільних сигналів з нескінченного набору, але коли у нас є набір дискретних дозволених станів діяльності, досить багато.
Тобто, виходить, що значення структури аксональних і дендрітичних дерев, властивих мозку, є створення на дендротичної поверхні кожного нейрона насиченого простору пасток, тобто місць, що відповідають всіляким поєднанням джерел нейротрансмітаторів. За щільністю нейротрансміттера в пастках можна судити просторово розподілений сигнал, що складається з синхронної активності ряду сусідніх нейронів.
І так звані метаботропні рецептори на поверхні нейрона, які можуть викликати одиночні адгезії нейрона навіть при невеликій концентрації нейротрансмітерів, виходить, що з цими адгезивами, які зазвичай називають спонтанними, нейрон може реагувати на величезну кількість різних сигналів, відрізняється від тих, на яких нейронних сипсів можна налаштувати.
Власне, тепер можна пояснити призначення тих самих дендротичних хребтів, з якими почалася стаття. В теорії всі сипси можуть розташовуватися безпосередньо на дендриті або тілі нейрона. Це не впливає на пряме функціонування сипсів і їх здатність сприяти індукованої активності. Але тепер ми можемо припустити, що мета дендритичних хребтів є створення просторової структури, в якій синопси різних нейронів є «змішаними», щоб вони придбували можливість впливати не тільки на поверхню власної, але і навколишні сусідні дендрити. Це «прискорена дрібниця». й
У оригінальній статті показано, як все це народжується правильним «хвильовим оповіданням», але це окрема розмова.
І, нарешті, маленька самозахідна. Оригінальна стаття в оригінальній Росії. Є переклад англійською мовою, виконаною Дмитром Шабановим (для якого він має велику повагу), але це далеко від ідеального. Якщо у кого-небудь є можливість шукати і відмітити відбій, я буду дуже вдячний. Відкрито документ для кожного коментаря.
Олексій Редутов
Перелік посилань на оригінальну статтю
1. (2014). Джерело: Human Connectome Project: www.humanconnectomeproject.org
2. (2014). AllEN Mouse Brain Connectivity Atlas: підключення.brain-map.org
3. Bloom, B.H. (1970). Космічні / часові торгові марки в хеш-кодуванні з допустимими помилками Комунікації АКМ Т. 13 (7), 422-426.
4. Брайтенберг В., Шуз А. (1998). Cortex: статистика та геометрія з’єднання нейронал, 2-е ед.
5. Умань Брайтенберг, В. (1978). Кортикальні архітектоніки: загальний і В М.А. Брайзер та Х. Петше (ред.), Архітектоніки Церебрального Cortex (пп. 443–465). Нью-Йорк: Равен Прес.
6. Костюм, Х. (1975). Електромеханічні напруження і ефект рН на мембранну структуру. Biochim Biophys Acta 13;382(2), 142-146.
7. Про нас Христен М. Харріс. Synapse Web. Доступ до synapses.clm.utexas.edu/.
8. Неомагніт Самоорганізована модель нейромереж для механізму розпізнавання шаблонів, неафілійованих за допомогою зсуву в положенні. Біологічна кібернетика, 36(4), 193-202.
9. Навігація Валенсберг, С. (1987). Конкурентне навчання: Від інтерактивної активації до адаптивного резонансу. Когнітивні науки N11, 23-63.
10. Геб, Д. (1949). Організація поведінця. Нью-Йорк: Джон Wiley & Sons.
11. Ходгкін, А. а. (1952). кількісний опис мембранного струму і його застосування для проведення і збудження в нерві. Фізіо Л. 117, 500-544.
12. Іжікевич, Є. М. (2007). Динамічні системи в нейронауці: геометрія збудливості та буріння. Лондон: MIT Прес.
13. Куллманн, Д. М. (2000). Суловер і симпатичний хрест розмови з глутаматом і ГАБА в мозку ссавців. Prog Brain Res, 125, 339-351.
14. Курамото, Ю. (1984). Хімічні осади, хвилі та турбулентність. Пувер Публікації.
15.00 р. Лібке Й., Маркрам Г., Францер М., Сакманн Б. (1996). Частота і дендротичний розподільчих прикусів, встановлених шаром 5 пірамідальних нейронів у розвитку щурів неокортекс: порівняння з синоптичним інертуванням сусідніх нейронів одного класу. Неврсі 16 (10), 3209-3218.
16. Маленка Р.К., Нікол Р.А. (1999 р.). Довгострокова потенціація-за десятиріччя прогресу? Наука 285(5435), 1870-1874.
17.00 р. Михаїл Т. Ліпперт, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu. (2007) Методи напруга-чутливого Dye Imaging of Rat Cortical активність з. J Neurophysiol 98, 502-512.
18. Піттс В., Маккулох В.С. (1947). Як ми знаємо універсальності: сприйняття слуху та візуальних форм. Бул. Маф. Біофіз В.9, 127-147.
19. Розенблат, Ф. (1962). Принципи нейродинаміки: Перцепрони та теорія механізмів мозку
20. Русаков Д. А., Куллман Д. М. (1998). Додаткові синоптичні глутаматні дифузії в hippocampus: ультраструктурні обмеження, поглинання і активація рецептора. Неврсі 18(9), 3158-3170.
21. Шен, М., Накагава, Т. (2002). Нейробологія: рецептори глутамату на ходу. Природа, 417(6889), 601-602.
22. Шен, М., Сала С. (2001). домени PDZ і організація супромолекулярних комплексів. Анну Рев Нейросі, 24, 1-29.
23. Сикова Є., Мазель Т., Вагрова Л., Ворік І., Прокопова-Кубінова С. (2000). Додатковий простір дифузії та патологічних станів. Прогрес в Brain Research, 155-178.
24. Товар К. Р., Вестброк Г. Л. (2002 р.). Мобільні рецептори NMDA на стегнах нейрон, 34(2), 255-264.
25. В.-Ф. Су, X.-Y. Гуан, К. Такагаки, Ю.-Ю. У. (2007). Пригнічення і відбиття візуально провокованих кортичних хвиль. Нейрон, 55, 119-129.
26. Ю. Лекун, Ю. Беніо (1995). Конволюційні мережі для образів, мови та часових серіалів, в Арбіб, М.А., редактор, Handbook of Brain Theory and Neural Networks. МИТ Преса.
27. С.П. Шнурова, З.М. Гвоздикова. (1971). Реакція нейронів в корі сенсорів до безпосередньої електричності. У збірнику «Дослідження організації нейронної активності кори великих півсфер головного мозку» (пп. 158-180). Москва: Наука.
28. Нихеллс Ю., Мартин Р., Валас Б., Фукс П. (2003). Від Nero до Brain (Fourth Edition)
29. Я очищаю, К. (1971). Біблії мови.
30. Радченко А.Н. (2007). Аналізи інформації про мозок. Сб.
31. Редозув, А. (2014). Програма обчислення розподілу сиптичних пасток. Отриманий з aboutbrain.ru: www.aboutbrain.ru/programs/
32. Редозув, А. (2014). Програма моделювання хвильової кори. Отриманий з aboutbrain.ru: www.aboutbrain.ru/programs/
Джерело: habrahabr.ru/post/230047/