1642
Логіка мислення. Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Ця серія статей описує модель хвилі мозку, що істотно відрізняється від традиційних моделей. Я дуже рекомендую, що ті, хто тільки приєднався до читання першої частини.
Поговоримо про нейронні детектори. Припустимо, що деяка інформація проводиться до критичної зони через хвильові тунелі. Кожна з проекційних променів – осьові нейрони, що розташовані на зоні, що надсилає цю інформацію. Проекція знімається з невеликої площі кори. Волокно проекційні балки, фактично, трансляційні хвилі, що проходять через цей сайт. Місце проведення проекції стає джерелом хвиль. Ці хвилі здійснюють однакову інформацію в зоні отримання скоринки як хвилі в оригінальній зоні.
Якщо ми налаштуємо ваги нейрона, щоб розпізнати певну хвилю, що проходить через її рецепційне поле, ми перетворюємо її в детектор, який вогнегасає при його характерному поєднанні ідентифікаторів.
Якщо ми навчаємо кількох нейронів, розташованих у відносній близькості один до одного, щоб виявити однаковий візерунок хвилі, ми не маємо єдиного нейронного детектора, але шаблон детектора. Відповідність одного нейронного детектора до характерного стимулу - це пакет імпульсів провокованої активності. Реакція шаблону детектора - індукована активність групи нейронів, які утворюють конкретний візерунок. Природно, такий візерунок провокованої активності почне навчати кору, щоб пропагувати свій унікальний ідентифікатор.
Виявляється, коли кортична зона отримує опис, що складається з певних функцій, її шаблони детекторів відповідають знайомим поєднанням і генерують власні хвилі, які здійснюють інформацію про нові функції, визначені цією зоною. Таким чином, корова зона вперше виявляє статистичні закономірності на зображенні, що надходить до неї за допомогою функції та форм узорів-детекторів, що відповідають цим нормам, а потім, отримання описів в деяких рисах, генерує власний опис у тих рисах, які були виявлені раніше цією зоною.
Логіка цієї трансформації нейро описів можна порівняти з логікою словесних описів, що не дивно, оскільки ці явища тісно пов'язані. Ми бачимо різні тварини. Ми можемо описати кожен з них набором знаків. Наприклад, ми бачимо звір з описом: великий, коричневий, shaggy. Не знаючи, який вид здоби його, ми можемо назвати його, що: великий коричневий шашлик. Поєднання характеристик формує ідентичність цього тварини. Якщо цей звір виникає нам часто, то ми можемо виправити закономірність того, що ці ознаки відбуваються разом, і дати це поєднання власної назви, наприклад, ведмедика. Далі ми можемо використовувати термін «паш» як самостійну концепцію. У той же час у нас є можливість, якщо необхідно, щоб відновити всі початкові ознаки терміном «задній».
Висловлення реального світу, зручно використовувати термін «феномон». Його зручність в громаді. Цей термін описує не тільки об'єкти, але і процеси, що відбуваються з ними. Ми можемо викликати будь-яку картину, яку ми сприймаємо феномен. Кожне явище ми генеруємо опис. Опис – сумарність роботи детекторних нейронів, що утворюють візерунки та ідентифікаційні хвилі, що створюються ними.
Ви можете викликати шаблон нейронних детекторів, які вогонь на одному зображенні та відповідну концепцію ідентифікації хвилі. Тоді ми можемо сказати, що будь-яке явище представлена мозку як набір описів, що складаються з концепцій. Для кожної зони кори описується власним набором концепцій.
При детекторі нейронів випромінюють новий візерунок або закріплюємо нове поєднання особливостей, щоб утворився візерунок детекторів, що може генерувати свою унікальну хвилю, ми можемо говорити про формування нової концепції. Розшифровка цієї концепції може бути справжніми світовими подіями, які створили це поєднання рис, і до яких відповідні нейрони тепер відповідають.
Формування візерунка. Зворотна проекція
Візерунок детектора нейронів, спрямованих на загальний подразник, дозволяє викликати власну унікальну хвилю ID. Це відповідає розпізнанню образу і в тому числі концепції консультування в описі чого відбувається. Для ідентифікатора бути унікальним, достатньо для детектора нейронів, щоб створити візерунок на відміну від будь-яких хвильових візерунків, що проходять через їх розташування. Розкажіть просту модель, яка створює такий візерунок.
Повернемося до опису механізмів нейронної взаємодії. Головний спосіб передачі сигналів з одного нейрона в інший - випуск нейротрансмітаторів з аксонного терміналу передачі нейрона в симпатичну клітку. На поверхні нейрона, що отримує сигнал, є рецептори, які відповідають певному нейротрансмітеру, що є лігандом для нього. Деякі рецептори розташовані в симпатичних засобах і відповідають за симпатичну передачу, деякі зовні симпатики і відповідають за активність хвиль.
Є багато нейротрансмітерів, в тому числі нейроактивних пептидів, є більше п'ятдесят з них. Найпоширеніший активуючий нейротрансміттер – глутамат, пригнічений – гамма-амінобутирова кислота (ГАБА). Уміння активувати або пригнічувати не майно самого нейротрансміттера, але визначається типом рецептора, що взаємодіє з ним. Для передачі сигналу необхідно, щоб нейротрансмітент, який випромінив аксон і рецептори, які отримують його, відповідають один одному.
До кінця 80-х років минулого століття, вважається, що кожен нейрон видає той самий медіатор в усіх терміналах його осьового принципу. Але згодом показано, що є нейрони, які використовують різні комбінації медіаторів.
Візьміть два гіпотетичні типи нейронів, які працюють з одним передавачем, зателефонуйте його (A). Відмінність типів нейронів буде де вони будуть чутливі до загального медіатора (табл. нижче).
Синапс
Зовні симпатики
аксон
Тип 1 - А
Тип 2 А - Нейротрансміттер Характеристики нейронів. Чутливість рецептора і едикція аксона
Нейрони першого типу не мають рецепторів у своїх синапсах, які чутливі до медіатора. Але вони мають ці рецептори на екстрасинаптичну частину мембрани. Це означає, що ці нейрони не здатні до синоптичного навчання на візерунках активності, які створюються нейронами як перших, так і других типів, але здатні навчитися пропагувати активність хвиль і бути передавачами хвилі ідентифікації.
Тепер візьміть кору, яка складається з нейронів обох типів. Наприклад, просто перемішайте їх випадковим чином. Наявність інтерсперсованих нейронів другого типу не вплине на здатність кори вчитися проводити хвилі ідентифікаторів за допомогою нейронів першого типу.
Ми хочемо, щоб створити шаблон детектора.
Подати заявку на навчання
Повідомляємо, що ми вирішимо всі питання навчання, і наша робота зараз полягає в тому, щоб просто виправити прототип. Ми вже знаємо. Давайте отримаємо другий тип нейронів, які знаходяться в цій області і не займаємося навчанням, брати участь у генерації випадкових походів. Встановіть ймовірність адгезії, щоб виготовити якомога більше активних нейронів як нейронів детектора, нам потрібно в кінцевому шаблоні. Випадкові адгезії створять унікальний візерунок нейронів другого типу, локалізованих в обраній області.
Далі, відповідно до принципу Hebb, ми використовуємо активність нейронів другого типу як сигнал для їх навчання. Зміна ваги їх сипсів за допомогою хвильового паттерна вони дотримуються їх рецептивних полів. Результатом є закономірність нейронів детектора, спрямованих на новостворений набір ідентифікаторів. Відповідність хвильового візерунка призведе до передбачуваної активації всіх нейронів у шаблоні. Нейрони першого типу підійдуть цей візерунок і перетворять його в поширення хвилі нового ідентифікатора.
Ми отримали дуже простий і чудовий результат. Особливе поєднання ідентифікаторів може генерувати свій унікальний ідентифікатор.
Тепер робіть модель трохи складною. Додати інший медіатор (B) і змінити характеристики нейронів (табл. нижче). Синапс
Зовні симпатики
аксон
Тип 1 Б
Р
Тип 2 A B Характеристики нейронів з використанням двох посередників. Чутливість рецептора і едикція аксона
Поведінка такого дизайну буде набагато цікавіше. Нейрони другого типу, створивши шаблон детектора, не вдасться пропагувати хвилю від неї через нейрони першого типу, так як їх тип аксона медіатора не збігається з додатковим синоптичним чутливістю нейронів першого типу. Але ці нейрони зможуть пропагувати хвиля свого стилю. Цікаво, що відокремиться вхідна і вихідна активність кори. Якщо ми створюємо вихідний хвильовий тунель з осей нейронів другого типу десь на боці, то він не передає весь хвильовий візерунок, який складається з інших речей, безлічі функцій введення, але буде передавати лише опис, що складається з визнаних концепцій.
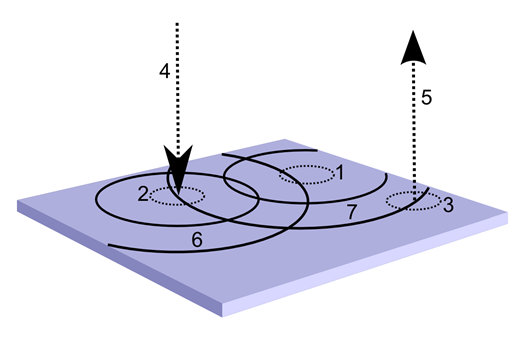
Зміна коретичного опису. 1 – вибрана робоча зона, 2 – область, виділяючи ідентифікатор вводу, 3 – область виходу, 4 – вхідної хвилі тунелю, 5 – вихідний канал, 6 – вхідна хвиля ідентифікатора, 7 – вихідна хвиля ідентифікатора
Але це не все. У тій же обраній області ми можемо виконати синоптичну підготовку нейронів першого типу. Для них сигнал, який навчиться, буде власною хвилею. Оскільки ми дали чутливість синхронів першого типу тільки до медіатора (B), вони сприймають малюнок того ж візерунка, який ми раніше навчалися на нейронах другого типу. У той же час, детектор нейронів першого типу буде підкреслюватися в шаблоні, що повторює хвилю ідентифікаторів, які відбувалися в момент тренування. Це означає, що якщо ми повертаємо на кору через вхідний хвильовий тунель, ідентифікатор поняття, який раніше відкриваються нейронами другого типу, потім поширюється на вибране місце, це призведе до активності шаблону детекторів нейронів першого типу. І так як ці нейрони повторюють візерунок фрагмента хвилі оригінальних ідентифікаторів, їх виклична активність призведе до відповідних ідентифікаційних хвиль, щоб поширюватися через нейрони першого типу.
Так ми відтворили зворотний проекційний механізм. Кора в нашій моделі здатна передавати інформацію в обох напрямках. У прямій спрямованості інтегровані функції і опис змінено на більш загальний. У зворотному напрямку отримав відповідний ідентифікатор, кора відновлює всі відповідні ознаки і переводить їх вниз, якщо, звичайно, є відповідні відносини проекції.
Ми отримали досить простий дизайн: в одному напрямку – навчимося концепту, в протилежному напрямку – відреставруємо знаки. Але це простота двостороннього зв'язку унікальна для нейромереж. Традиційні мережі динамічно непрямі, вони не можуть запускати сигнал “задній віссон” і цей недолік не лікується будь-якими хитрощами. У той же час, очевидно, що реальний мозок здатний не тільки узагальнити інформацію, підняти опис від рівня до рівня, роблячи його більш абстрактним, але і перевести опис назад, перетворюючи його на зображення, внутрішню мову, фізичні дії. Таким чином, це особливо те, що наш дизайн дозволяє нам відтворювати зворотну проекцію досить природно, зберігаючи в рамках механізмів, доступних для реального мозку.
Якщо ви робите осі нейронів одного типу генеруємо два медіатори одночасно, то ми отримуємо розподіл відповідних ідентифікаторів через два шари одночасно. Ви можете уявити ситуації, де це буде корисним. В цілому поєднання медіаторів і рецепторів можуть запрограмувати найбільш несподівану поведінку кори. У той же час, приємно, що наша кора не вимагає особливого топологічного регулювання. Нейрони утворюють синоптичні контакти з їх оточенням, не турбуючись про наступні будь-які схеми. Здається, що подібні архітектурні принципи притаманні реальній скоринки.
Література використана
Продовження
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Частина 5. Брайн хвилі
Частина 6. Система проекції
Частина 7. Інтерфейс користувача
Частина 8. Ізоляція чинників в хвилях мереж
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/215283/

















Логіка мислення. Зареєструватися 8. Ізоляція чинників в хвилях
Логіка мислення. Зареєструватися 10. Просторова самоорганізація