3163
Логіка мислення. Зареєструватися 10. Просторова самоорганізація

Ця серія статей описує модель хвилі мозку, що істотно відрізняється від традиційних моделей. Я дуже рекомендую, що ті, хто тільки приєднався до читання першої частини.
Ми приступаємо до того, що явища зовнішнього світу впливають на наші почуття, викликаючи певні потоки сигналів в нервових клітинах. У процесі навчання кора набуває можливості виявити певні комбінації сигналів. Детектори – нейрони, синоптичні ваги яких налаштовуються до закономірностей активності, що відповідають явищам, виявлених. Кортичні нейрони відстежують місцеве середовище, яке формує місцеве рецепційне поле. Відомості про рецептивні поля нейронів набувають як через топографічну проекцію, так і через пропагацію хвиль ідентифікаторів, що мають унікальні візерунки, що відповідають вже визначеним характеристикам. Детективні нейрони, які відповідають однаковому поєднанню схем детектора рис. Візерунки цих шаблонів визначають унікальні хвилі ідентифікаторів, які запускають ці візерунки, коли вони вводять стан провокованої активності.
У 1952 році Алан Турінг опублікував паперовий матеріал «Хіководія Морфогенезу» (Турінг А. М., 1952), присвячений самоорганізації речовини. Основним принципом він сформульовано, що глобальне замовлення визначається локальною взаємодією. Для того, щоб отримати структурну організацію всієї системи, не потрібно мати глобальний план, але можна обмежити себе виключно для встановлення правил тісної взаємодії елементів, що утворюють систему.
Вивчення нейронів не проводиться в ізоляції, але з урахуванням діяльності їх оточень. Правила ведення бухгалтерського обліку за цією діяльністю визначають самоорганізацію кори. Самоорганізація означає, що в ході навчання не просто виявляють нервові візерунки, але ці візерунки вбудовані в просторові конструкції, які мають певне значення.
Найбільш очевидним способом самоорганізації є структурування за безпосередністю. Можлива поїздка нейронів, щоб візерунки, що відповідають поняттям, які знаходяться поблизу і в космосі. Пізніше ми побачимо, що це розташування буде необхідно для багатьох важливих функцій мозку. У той же час ми розглянемо механізми, які можуть надати таку організацію.
А досить неоднозначним питанням є те, як вимірювати близькість поняття. Один підхід полягає в тому, що будь-яка концепція можна порівняти з описом. Наприклад, опис події може бути вектором, компоненти якого демонструють тяжкість певних особливостей події. Комплект функцій, в яких здійснюється опис дескриптивної бази. У цьому випадку розумний захід знаходиться поруч з описом. Чим ближче опис двох явищ, тим більше схожі ці явища. В залежності від проблеми, яка додатково вирішується за допомогою вимірювання відстані, можна вибрати один або інший алгоритм розрахунку. Запобігання близькій відстані між об'єктами. Можна розраховувати на інший і навпаки.
Ще один підхід – тимчасова близькість явищ. Якщо два явища відбуваються одночасно, але мають різні описи, то можна ще розповісти про їх певну близькість. Найчастіші явища відбуваються разом, чим більше їх близькість можна інтерпретувати.
По суті, ці два підходи не суперечать, але доповнюються. Якщо два явища часто виникають разом, то необхідно зробити висновок, що є явище, яке є втіленням цього поєднання. Це нове явище може бути ознакою як початкових явищ. Зараз ці явища мають загальний опис, а отже, афінність в дескриптивному підході. Дозвоніть, будь ласка, зв'яжіться з нами, вкажіть як дескриптивну, так і часову близькість.
Уявіть, що ви шукаєте щось з різних куточків. Можливо, ці види будуть досить різні в дескриптивних умовах, але незважаючи на їх близькість в іншому сенсі не можна відмовитися. Тимчасова інтимація - це можливість прояву цього різного значення. Трохи пізніше ми підемо набагато більш глибоким в цю тему, але зараз ми просто відзначимо, що необхідність врахування обох підходів при визначенні близькості концепцій.
Найлегша нейромережа для розуміння, яка ілюструє самоорганізацію за ступенем дескриптивної близькості, є самоорганізацією карт Коконен (рис. нижче).
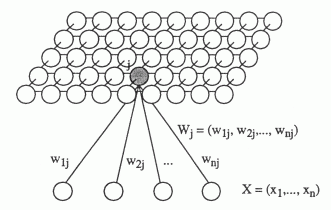
Давайте нам вхід, наведений вектором. Існує двовимірна сітка нейронів. Кожен нейрон підключений до вектора введення, це з'єднання визначається набором ваги. По-перше, ми ініціюваємо мережу з випадковими невеликими масштабами. Надаючи вхідний сигнал, для кожного нейрона, можна визначити його рівень активності як лінійний додаток. Візьміть нейрон, який показує найбільшу активність і викликав його нейрон. Потім перемістіть свою вагу до образу, він виглядав як. Ми зробимо все для всіх наших сусідів. Ми збираємось ослабити цей зсув, як ми відступаємо від нейрона-переможця.
Ось курс навчання, який знижується з часом, амплітуда верхологічного мікрорайону (в залежності від n припускає, що він також зменшується з часом).
Амплітуда мікрорайону можна вибрати, наприклад, за допомогою функції Gaussian:
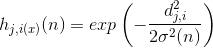
де знаходиться відстань між виправленою нейронною і нейронною перемогою.
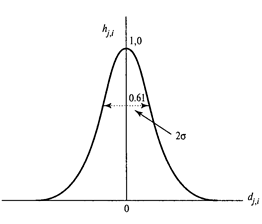
Функція Gauss
Як ви навчитеся, така самоорганізація карта висвітить ділянки, які відповідають яким чином розподіляються зображення. Це, сама мережа визначить, коли подібні фотографії зустрінеться в вхідному струмі і створять тісні представництва на мапі для них. У той же час, чим відрізняються зображення, тим більше виділяються один від одного їх представництва. В результаті, якщо ви забарвлюєте результат тренування відповідно, він буде виглядати приблизно так, як показано на малюнку нижче.

Результати навчання коханової картки
Панельні карти використовують функцію амплітуди верхнього мікрорайону, що говорить про те, що нейрони, крім взаємодії за допомогою симпозів, можуть обмінюватися додатковою інформацією про характер навколишньої активності, і ця інформація може вплинути на перебіг їх сиптичного навчання. Необхідність додаткової інформації виникла дещо раніше в нашій моделі. Зняти підготовку екстрасинаптичних рецепторів, ми ввели правила на основі знань нейронів, що оточують активність певного типу. Наприклад, знаючи загальний рівень діяльності дозволило нам приймати рішення про те, чи варто вчитися.
Для того, щоб залишатися в рамках певної біологічної певненості, ми спробуємо показати, які механізми в реальному корі можуть бути відповідальні за обчислення та передачі додаткової інформації, яка не закодована в сигналах axon.
Близько 40 відсотків мозку займають гліальні клітини. Загальна кількість про порядок більшої кількості нейронів. Традиційно гліальні клітини призначають безліч функцій сервісу. Вони створюють об'ємну раму, заповнюючи простір між нейронами. Зареєструватися в підтримці гомеостазу навколишнього середовища. У розвитку нервової системи беруть участь у формуванні топології мозку. Шванн клітини і олігодендроцити відповідають за мієлітацію великих осей, що призводить до багаторазового прискорення передачі нервових імпульсів.
Оскільки гліальні клітини не генерують потенціали дії, вони не безпосередньо залучені до інформаційної взаємодії. Але це не означає, що вони повністю позбавлені інформаційних функцій. Наприклад, плазмі астроцити розташовані в сірій матерії і мають численні високо розгалужені процеси. Ці процеси стимулюють навколишні симпатики і впливають на їх роботу (рис. нижче).
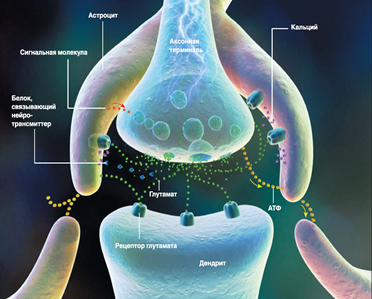
Астроцит і Синапс (Fields, 2004)
Наприклад, описано наступний механізм (Р. Д. Поля, Б. Стівенс-Грахем, 2002). Активація нейрона призводить до виходу молекул ATP від її аксона. ATP (аденозин трифосфат) - це нуклеотид, який грає надзвичайно важливу роль у всьому тілі, його основною функцією є забезпечення енергетичних процесів. Але крім того, ATP може діяти як речовина сигналізації. Під його впливом розпочато рух кальцію всередині астроциту. Це, в свою чергу, викликає астроцит, щоб звільнити свій власний ATP. В результаті цього стану передається на сусідні астроцити, які передають його ще далі. У цьому випадку засвоєння кальцію астроцитом призводить до того, що він починає впливати на сипси, з якими він контактує. Астроцити здатні одночасно посилити реакцію на сніпсування шляхом введення відповідного посередника, а також ослаблення його шляхом поглинання або видалення в’яжучих білків нейротрансміттера. Крім того, астроцити здатні звільнити молекули сигналізації, які регулюють випуск посередника за допомогою аксону. Концепція сигналізації між нейронами, яка враховує вплив астроцитів, називається триартитним сипсом.
Ця взаємодія астроцитів і нейронів не передає специфічні інформаційні зображення, але дуже добре підійде до ролі механізму поширення «бездіяльності поля», що дозволяє контролювати вивчення симпозів і тим самим встановлювати просторові координати для нових моделей детекторів.
Крім астроцитів, поведінка сипсів також впливає на міжклітинну матрицю. Матриця - це сукупність молекул, що виробляються клітинами мозку, які заповнюють міжклітинний простір. У статті (Дитатв А., Шахнер М., Синдерегер П., 2010) показано, що зміна складу матриці впливає на характер сиптичного пластичності, тобто вивчення нейронів.
Імпульсна активність нейронів створює точковий просторовий візерунок, що динамічно змінюється, кодування інформаційних потоків. Ця активність змінює стан гліального середовища та матричному середовищі таким чином, що створюється щось схоже на узагальнене поле діяльності (рис. нижче).

Діяльність та поле діяльності
Поле діяльності, з одного боку, відбійна точна активність, створення регіону, що поширюється за межі активних нейронів, з іншого боку, має інерцію і продовжує існувати протягом деякого часу після припинення імпульсної діяльності.
Якщо в межах цієї сфери діяльності ми створюємо детектор поточного подано зображення, він буде близьким до аналогічних детекторів. При цьому схожість може бути одночасно сприйнятливою схожістю і схожістю, що виникає з поєднання подій з часом.
У реальному мозку найбільша просторова організація для первинної візуальної кори. У зв'язку з трансформаціями, які починаються в сітківці, сигнал отримується на первинній кори, в якій основною інформацією є лінії, що описують контури об'єктів в оригінальному образі. Нейрони в первинній візуальній корі дивляться переважно невеликі фрагменти цих ліній, що проходять через їх рецептивні поля. Неприпустимо, значна частина нейронів в цій зоні є лінійними детекторами, які виконуються під різними кутами.
Експериментально, нейрони розміщують вертикально нижче одного відгуку один до одного. Група таких нейронів називається кортичним міні стовпчиком. Vernon Mountcastle (W. Mountcastle, J. Edelman, 1981) гіпотем, який для мозку, кортичний стовп є базовим структурним підрозділом обробки інформації.
У минулому, коли ми говорили про візерунки нейронів детектора, ми зображували їх як групи нейронів, розподілених в місцевій області. Це пов'язано з тим, що в моделюванні використані плоскі нейромережі, і відповідно, при підготовці картин. Реальна кора об'ємна. Об'єм кори не впливає на нашу причину виникнення і поширення хвиль ідентифікаторів. У тривимірній корі, пропагують хвилі таким же чином, як в квартирі. Але в об'ємній корі нічого не заважає нам від розміщення детекторів нейронів, які утворюють один візерунок вертикально нижче один одного. Ця композиція не гірше, ніж будь-який інший. Основною вимогою до шаблону є випадковість його шаблону. Оскільки з'єднання нейронів розподілені випадковим чином, нейрони, розташовані вертикально в одному з кортичних стовпчиків можна вважати випадковим малюнком. Це вертикальне розташування досить зручно при визначенні розташування шаблону виявлення. Не потрібно визначити місцеву площу, але не потрібно вказати позицію, в якій потрібно створити візерунок. Вибір положення може бути, наприклад, де поле діяльності максимальна для всіх вільних стовпців певного мікрорайону. Ми можемо припустити, що кортичні мікроколони реальної кори є шаблонами детектора, описаними в нашій моделі.
Для традиційних моделей кори, які не враховують хвильові сигнали, пояснюючи, що всі нейрони в мінікулах відповідають тим же стимулам, представляє певну складність. Ми повинні припустити, що це або небажання, щоб забезпечити толерантність до несправностей, або спробу об'єднати нейрони, які відповідають одному стимулу, але налаштовані для визначення його в різних положеннях загального рецептивного поля. Останні з'явився в неоконтейтроні через використання площин простих клітин. Модель хвилі дозволяє виглядати на ньому з іншого кута. Можна припустити, що завдання кортичного міні-ролика, дізнавшись характерний стимул, щоб запустити відповідний ідентифікатор хвилі. Потім кілька десятків нейронів, що утворюють мінікул і реагують разом – це механізм спуску хвилі, тобто шаблон детектора.
Організація орієнтаційних колон в реальному візуальному корі формує так звану «ручку». Такий «топ» (рис нижче (B) має центр, де колони різних орієнтацій конверж, і дивергентні хвости, які характеризуються гладкою зміною бажаного стимулу. Один топ формує гіперкул. Хвости різних гіперколомів переходять в один одного, утворюючи орієнтацію карти кори (рис. нижче (А)).

Розподіл орієнтаційних колон в реальному корі, отриманих методом оптичного (Ніхольс J., Мартін Р., Валас Б., Фукс П., 2003)
На прикладі візуальної кори зручно порівняти реальний розподіл орієнтаційних колон і результати моделювання за допомогою сфери діяльності.
Ми подаємо на фрагмент лінії кори під різні кути. Діяльність детектора нейронів буде розглядатися як козин кута між представленим зображенням і зображенням на детекторі.
Індексований нейрон має координати на кори. Вартість поля діяльності з нейрона в точці з координатами може бути представлена, наприклад, через розподіл Gaussian:
3610Р. 4200Р.
На кожній точці кори можна писати загальну вартість поля діяльності:
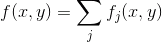
Ми розміщуємо нові детектори в безкоштовному положенні, для якого максимальна вартість. В результаті такої підготовки показано на малюнку нижче.

Результат навчання детектора (ліворуч), поле діяльності (праворуч)
Як бачимо, результат моделювання дуже схожий на реальний розподіл. Але слід зазначити, що на прикладі дуже умовний. У справжню кору кожен стовпчик злегка зміщується відносно своїх сусідів і слідує злегка різному фрагменту зображення. Це створює хвостики детекторів, що налаштовують на одну спрямованість. Але стимули цих детекторів відрізняються зображеннями на основі різних приймальних полів.
Просторова самоорганізація кори можна порівняти з самоорганізацією речовини у всьому світі. Атоми, молекули, об'єкти, планети, зірки, галактики, Всесвіт – це всі наслідки існування чотирьох фундаментальних взаємодій. Ми можемо самі зателефонувати одержувачу і узгодити зручний час. Часті питання створюють поля навколо них, що відповідають взаємодіям. Схожі поля підведені і утворюють отримане поле. Отримані поля впливають на частинки, визначаючи їх поведінку. Те ж саме відбувається з мозку. Здається, що скоринка може створити кілька видів полів з різними властивостями. Кожне поле має свій вплив на поведінку нейронів. Поєднання цих взаємодій, наприклад, може визначити алгоритм синоптичного навчання і, таким чином, просторова організація нейронів детектора.
Легко побачити, що просторове угрупування в узагальненій близькості містить внутрішню суперечність. У зв'язку з цим я згадаю анекдоту. Ліон вирішив розділити тварин на красиві і розумні. І стоїть підкована мавпа: "І що я відірвати себе зараз?" Відрізки виникають при розміщенні концепції близько до інших, віддалених концепцій. Наприклад, якщо ми влаштовуємо книги на полицях бібліотеки, ми одночасно хочемо групувати їх жанрами, авторами та оригінальною мовою, а також рейтингом читачів. Якщо є тільки одна книга, є проблема.
У процесі самоорганізації кори може вийти те, що однакова концепція закривається в узагальненому сенсі на кілька інших, розташованих в різних місцях кори. Не має сенс розмістити шаблон детектора в компромісі, намагаючись знайти якесь рівноважне місце. По-перше, таке місце не може бути, а по-друге, повністю порушує принцип «сімейстер на наступний». Немає нічого лівого, але для створення декількох шаблонів детектора, кожен близько до локального максимального поля діяльності.
На перший погляд, думаність того ж поняття в різних місцях виглядає потворно. Робота з інформацією, ви завжди хочете приїхати до її інтеграції, не атомізації. Але наша модель нейромереж дозволяє нам елегантно видаляти протиріччя, які виникають. Кожна з шаблонів детектора, хоча створена в різних місцях в корі, навчається на одній хвилі ідентифікаторів. У той же час, якщо ви берете дворівневий дизайн, один з рівнів захоплює фрагмент хвильового малюнка загального ідентифікатора. Це означає, що шаблони детектора, що розсіяні в різних місцях, з'єднуються в одній концепції з загальним ідентифікатором.
Ця поведінка концепцій є наслідком раніше заявленого дуалізму. Концепція - як візерунок, так і хвиля. Візерунки генерують хвилі, хвилі активізують візерунки. Просторове положення візерунків, пов’язаних з однією концепцією, характеризує концепцію через її близькість до інших концепцій. Про властивості і особливості відповідного явища говорить позиція візерунки на корі, фактично, говорить багато про властивості і особливості відповідного явища. Але якщо одна область відповідає одному концепту на картах Коконена, так як вона виходить з парадигми «переможець забирає все», то наша модель управляється не тільки багатим описом, але і зберегти єдність компонентів через дуалізму.
Самоорганізація за принципом «символа на наступну» не є кінцем для кори. Після цього ми покажемо, що найважливіші алгоритми головного мозку вимагають такої організації і практично неможливо реалізувати зовні.
І тепер це не зовсім банальна фізична аналогія. Основна ідея квантової фізики полягає в тому, що квантова система не може бути в будь-якому стані, але тільки в деяких, допустимих для неї. У цьому випадку квантова система не займає певного значення, доки не відбувається вимірювання. В результаті вимірювань система переходить в одну з дозволених станів. Вимірювання будь-якої зовнішньої взаємодії, яка викликає квантову систему, щоб проявляти себе.
У квантовій фізики введено комплексно-цінну функцію, яка описує чистий стан об’єкта, який називається функцією хвилі. У найбільш поширеній інтерпретації Копенгагена ця функція пов'язана з ймовірністю виявлення об'єкта в одному з чистих станів (пл. модуля функції хвилі - щільність ймовірності). Система гамільтонської системи - це динамічна система, яка описує фізичні процеси без розсіювання. До квантової системи взаємодіє, це Гамільтоніан. Поведінка такої системи в часі і просторі можна описати описом еволюції її хвильової функції. Ця еволюція визначається рівнянням Schrödinger.
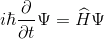 Р
Рде Гамільтоніан є загальним енергетичним оператором системи. Його спектр є набором можливих станів, в яких система може знайти себе після вимірювання.
Коли відбувається вимірювання, квантова система припускає одну з дозволених станів. Це називається зменшенням або руйнуванням функції хвилі. Під час процесу вимірювання система перестає бути ізольованою, а її енергія не може зберігатися, оскільки енергія обмінюється з пристроєм.
Який стан квантової системи припустимо в момент вимірювання – справа шансу. Але ймовірність вибору кожної з держав не така ж, вона визначається значенням функції хвилі, пов'язаної з цим станом.
У нашій моделі шаблони детекторів створюються в кортильних позиціях, де є дескриптивна або часова близькість нової концепції та концепції вже існуючої. Таким чином, створюється сукупність місць, що характеризують ймовірність прояву поняття в певній ситуації. Це можна порівняти з повноенергетичним гамільтонським спектром квантової системи. Пропагація хвильових ідентифікаторів можна порівняти з еволюцією функції хвилі. Опис, який ідентифікатор переходить в активність візерунків. Далі ми покажемо, що не всі візерунки активовані, але тільки ті, які можуть описати послідовну картину, відповідну контексту чого відбувається. У цій точці наша система переходить в одну з можливих дозволених станів, яка дуже схожа на злам хвильової функції.
Література використана
Продовження
Попередні частини:
Частина 1. нейрон
Зареєструватися 2. Фактори
Частина 3. Перцептрон, забруднені мережі
Частина 4. Підземна активність
Частина 5. Брайн хвилі
Частина 6. Система проекції
Частина 7. Інтерфейс користувача
Частина 8. Ізоляція чинників в хвилях мереж
Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Олексій Редозубов (2014)
Джерело: habrahabr.ru/post/215287/

















Логіка мислення. Зареєструватися 9. Нейрон-детекторні візерунки. Зворотній проекції
Логіка мислення. Зареєструватися 11. Динамічні нейромережі. Асоціативність