Жизнь — интересная!
Подписывайтесь на нашу группу в Telegram и Facebook, чтобы быть в сообществе единомышленников, находить вдохновение и не пропускать свежие и удивительные статьи с bashny.net.
1873
0.4
2014-03-21
Логика мышления. Часть 14. Гиппокамп
Этот цикл статей описывает волновую модель мозга, серьезно отличающуюся от традиционных моделей. Настоятельно рекомендую тем, кто только присоединился, начинать чтение с первой части.
Полное удаление гиппокампа делает невозможным формирование новых воспоминаний, что убедительно продемонстрировал случай с пациентом H.M. Нарушения в работе гиппокампа могут привести к синдрому Корсакова, который так же сводится к невозможности фиксировать текущие события, при сохранении старой памяти. Все это убеждает в том, что гиппокамп играет ключевую роль в механизме памяти.
Традиционные теории о роли гиппокампа строятся на аналогии мозга и компьютера. В таких рассуждениях гиппокампу отводится роль «оперативной памяти», то есть места, где накапливаются новые воспоминания. Затем же, предположительно во сне, эти воспоминания переносятся в области мозга, ответственные за хранение долговременной памяти. И хотя механизмы такого переноса непонятны, но, по крайней мере, это позволяет объяснить, почему нарушение работы гиппокампа блокирует формирование событийной памяти.
Наша модель принципиально отличается от моделей традиционных. У нас не происходит никакой буферизации памяти в гиппокампе и последующего ее копирования. Элементы, образующие воспоминания, сразу формируются там, где им уместно находиться. Гиппокамп же при этом просто создает единый идентификатор, который объединяет распределенные по пространству коры элементы памяти. Волновая модель работы мозга объясняет, как этот идентификатор распространяется по всей коре. Наличие такого идентификатора позволяет, выбрав объединенные им элементы, воспроизвести не абстрактную картину, а конкретный образ запомненного события.
В 2010 году были проделаны интересные опыты по протезированию гиппокампа крысы (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011). Подопытным животным с обеих сторон мозга в гиппокамп внедрялись микроэлектродные массивы, захватывающие области CA3 и CA1. После того, как крысы восстанавливались после операции, начинались серии тестов.
Крыс помещали в камеру с двумя выдвигающимися рычагами. Тест состоял из трех фаз. Сначала случайным образом выдвигался один из рычагов. Когда крыса нажимала этот рычаг, фиксировалась картина активности предварительно отобранных нейронов гиппокампа (стадия образца). Затем рычаг убирался, и наступала пауза от 1 до 30 секунд в одних сериях и до 60 в других. После чего выдвигались оба рычага. При нажатии на рычаг, противоположный исходному образцу, выдавалось подкрепление в виде капельки воды. При ошибке, то есть нажатии на тот же рычаг, что был на стадии образца, следовало наказание – выключение света на 5 секунд. В результате составлялся график зависимости успешности попыток от времени задержки (рисунок ниже).
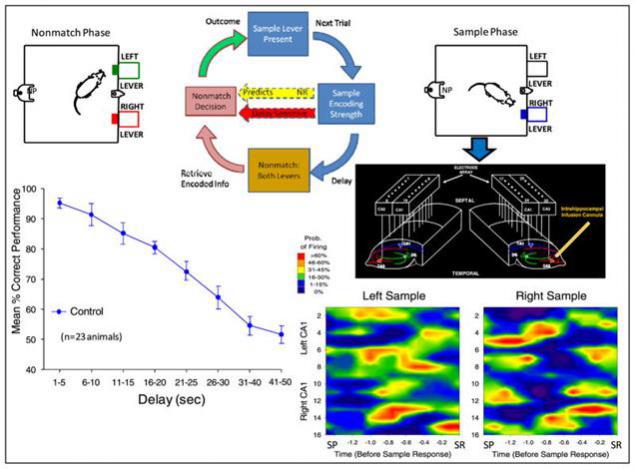
Контрольные измерения при естественной активности гиппокампа (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Исходя из записанных сигналов, была составлена предсказательная модель, которая прогнозировала, какой сигнал следует ждать на CA1 в зависимости от сигнала на CA3. В серии опытов сигналы прогноза были добавлены к существующим сигналам (рисунок ниже).
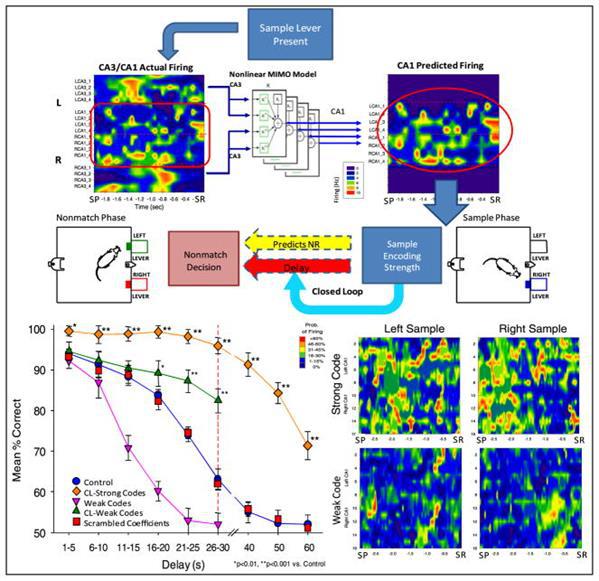
Добавление искусственных сигналов (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Получился интересный результат. Оказалось, что существенное влияние на результат оказывается только при корректировке сигнала в стадии предъявления образца, то есть при запоминании исходного образа. Картина же сигналов в остальные моменты была не принципиальна. Причем, чем сильнее был корректирующий сигнал, тем лучше был итоговый результат.
Авторы эксперимента, объясняя результат, исходили из представления, что гиппокамп занимается кодированием текущей информации, подготавливая ее для записи в память. Что картины активности, наблюдаемые в гиппокампе, несут информационное описание происходящих событий. Соответственно, усиление «правильного» сигнала способствует улучшению результатов запоминания.
В нашей модели объяснение полученных результатов несколько иное. Понятно, что идентификатор гиппокампа важен только в момент формирования памяти. После того как воспоминание создано гиппокамп не требуется для его воспроизведения. Усиление текущего идентификатора гиппокампа положительно сказывается на силе запоминания, но это не результат создания более четкой информационной картины, а просто выделение текущего воспоминания на фоне остальной памяти. Если у нормально работающего мозга таким образом подчеркивать все воспоминания, то итоговый результат окажется нулевой.
Самая интересная серия опытов была связана с подавлением собственной активности гиппокампа. Для этого крысам делались регулярные инъекции в область CA3 гиппокампа препарата MK801. MK801 блокирует нормальную работу синаптической передачи, использующую глутамат. Он проникает в ионные каналы NMDA рецепторов, чувствительных к глутамату и нарушает их работу.
Блокада участка CA3 ожидаемо резко ухудшила результаты. Но подача искусственных сигналов, восстанавливающих ожидаемую картину активности, позволила значительно улучшить количество правильных ответов (рисунок ниже).
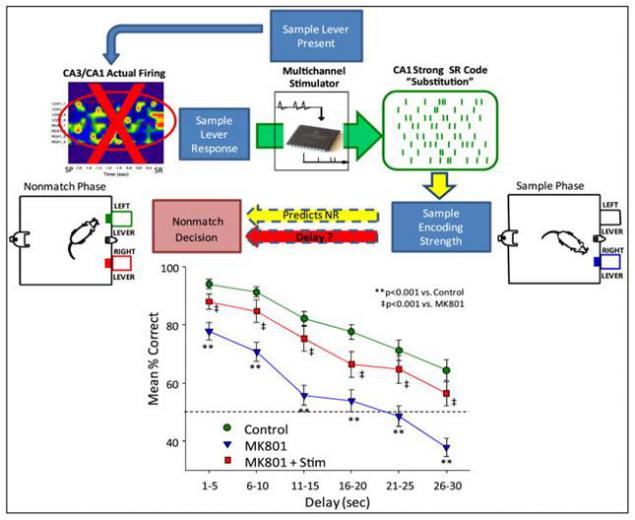
Замена естественного сигнала гиппокампа искусственным (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Я не склонен толковать этот результат как чудесное восстановление нейронного описания происходящего в гиппокампе с последующей записью этого описания в память. Тем более что в опыте использовались матрицы со всего 32 электродами, из которых задействовано было около половины. Скорее всего, происходило создание случайного идентификатора, который и делал возможным формирование памяти.
Но не все так просто с гиппокампом. Есть несколько фактов, которые достаточно сильно озадачивают при первом знакомстве с ними. Так в 1971 году Джон О'Киф открыл в гиппокампе клетки места (O'Keefe J., Dostrovsky J., 1971). Эти клетки реагируют подобно внутреннему навигатору. Если крысу поместить в длинный коридор, то по активности определенных клеток можно будет точно сказать, в каком его месте она находится. Причем реакция этих клеток не будет зависеть от того, как она попала в это место.
В 2005 году в гиппокампе были обнаружены нейроны, кодирующие положение в пространстве, образующие нечто наподобие координатной сетки (Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I., 2005).
В 2011 году оказалось, что в гиппокампе есть клетки, которые определенным образом кодируют временные интервалы. Их активность образует ритмические узоры, даже если ничего другого вокруг не происходит (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011).
Из всех этих фактов напрашивается вывод, что гиппокамп отвечает за кодирование нашего положения, как во времени, так и в пространстве. При этом гиппокамп подобен солдату, для которого вполне естественно «копать от забора и до обеда». Гиппокамп соединяет в себе и пространственную, и хронологическую навигацию, которые, кстати, имеют много общего. Так же, как наше путешествие в пространстве происходит от ориентира к ориентиру, так и путешествие во времени проходит от одной хронологической метки к другой.
Мы ранее утверждали, что гиппокамп формирует уникальные идентификаторы для воспоминаний. Как соотносится эта функция с тем, что гиппокамп отслеживает наши пространственно временные координаты? Конечно, можно считать, что гиппокамп несет двойную функцию, участвуя в двух независимых процессах. Но более логично предположить, что мы имеем дело с двумя сторонами одной медали.
Как мы идентифицируем информацию вообще? Используя Google, мы составляем запрос, состоящий из набора слов, который определяет смысл нашего поиска. Получив список результатов, мы затем можем уточнить его, введя ограничения по дате или по географии.
Создавая библиотеку изображений, мы помечаем фотографии датой их создания и геотегами места, где они сделаны. Потом мы описываем, кто или что запечатлено на них.
Размещая файлы на компьютере, мы указываем время создания и путь к файлу. Названием файла мы кратко описываем его содержание.
Получается, что идентифицируя самые разные вещи, мы используем, с одной стороны, признаки, описывающие их смысл, а, с другой стороны, координаты в пространстве и времени, как универсальные признаки любых явлений. Такая идентификация впоследствии оказывается достаточно удобной и полезной не только как уникальная метка, но и как инструмент поиска. Можно предположить, что и природа пошла аналогичным путем.
То есть, создавая уникальный идентификатор события, гиппокампу имеет смысл не просто генерировать случайный код и распространять его по всей коре, а закладывать в этот код пространственно-временные признаки, как наиболее подходящие для всех случаев жизни. Такое пространственно-временное описание само по себе оказывается достаточно уникально, как сочетание разнообразных факторов. Достаточно добавить к нему некий небольшой случайный довесок и получившийся идентификатор, будет однозначно указывать на конкретное событие.
Такая идентификация значительно мощнее по своим возможностям, чем просто уникальный ключ для каждого воспоминания. Появляется возможность разнообразных дополнительных ассоциаций. Но самое главное, появляется возможность кодирования сложных описаний, учитывающих то, как развиваются события и во времени, и в пространстве. Но разговор об этом будет несколько позже.
Если допустить, что идентификатор гиппокампа действительно кодирует пространственно-временные координаты, то получается, что гиппокамп, во-первых, обладает способностью взаимодействовать с корой таким образом, чтобы иметь возможность обучаться узнавать похожие позиции в пространстве и времени. Во-вторых, чтобы распространять свои идентификаторы по всему пространству коры, гиппокамп должен формировать их из конечного числа уникальных фрагментов. Это проистекает из того, что кора должна быть предварительно обучена на распространение всего «алфавита», используемого гиппокампом. Новые идентификаторы должны формироваться не «с нуля», а как комбинации уже известных элементов. В принципе, в этих требованиях нет ничего сложного, это соответствует тому, как в описаниях на естественном языке все фразы строятся из конечного набора букв.
Использованная литература
Продолжение
Предыдущие части:
Часть 1. Нейрон
Часть 2. Факторы
Часть 3. Персептрон, сверточные сети
Часть 4. Фоновая активность
Часть 5. Волны мозга
Часть 6. Система проекций
Часть 7. Интерфейс человек-компьютер
Часть 8. Выделение факторов в волновых сетях
Часть 9. Паттерны нейронов-детекторов. Обратная проекция
Часть 10. Пространственная самоорганизация
Часть 11. Динамические нейронные сети. Ассоциативность
Часть 12. Следы памяти
Часть 13. Ассоциативная память
Алексей Редозубов (2014)
Источник: habrahabr.ru/post/216409/
Портал БАШНЯ. Копирование, Перепечатка возможна при указании активной ссылки на данную страницу.