Жизнь — интересная!
Подписывайтесь на нашу группу в Telegram и Facebook, чтобы быть в сообществе единомышленников, находить вдохновение и не пропускать свежие и удивительные статьи с bashny.net.
634
0.2
2016-10-07
Структура и стартовые настройки мозга
Знание того как работает нейрон недостаточно для понимания того, чем обусловлено разумное и интеллектуальное поведение. Эволюция совершила удивительное мастерство, создав систему из относительно простых элементов, способную поразить невероятной успешностью во взаимодействии с окружающей средой.
Недостаточно взять некую массу связанных нейронов (даже сложив слоями) подключить к ней датчики и выводы и получить хоть какое-то подобие мозга. Главным полем работы эволюции на протяжении миллионов лет является не нейрон, а структура и внутренняя организация нервных клеток в нервной системе.

Давайте разберемся в том, как организована кора больших полушарий, структура благодаря которой Человек стал самым успешным видом на Земле.
Известно, что кора больших полушарий мозга человека состоит из шести условных слоев, но к такой форме она пришла за многие этапы эволюции. Поэтому для начала мы будем рассматривать упрощённый вариант с 2 – 3 слоями, ведь и такой вариант в природе имеет место. Любую биологическую структуру выгоднее изучать с позиции её эволюционного развития. Эволюция шла двумя путями развития коры: это увеличение количества слоёв и увеличение общей площади коры.
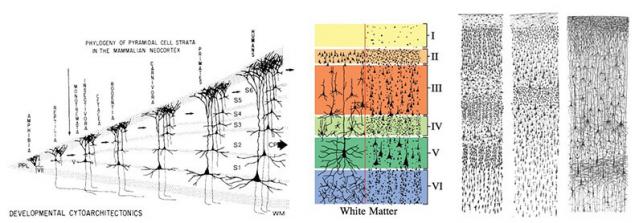
Второй особенностью коры является наличие так называемых кортикальных колонок. Можно сказать, что кортикальная колонка это логическая единица структуры коры. Рост самой коры во время эмбрионального развития осуществляется делением, самокопированием целых колонок. И это логично, если мы хотим сохранить некую структуру при росте.
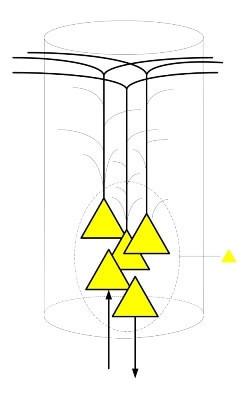
В основе кортикальных колонок лежат пирамидальные нейроны. Пирамидальные нейроны одни из самых крупных нейронов нервной системы, поэтому наиболее изученные, так как их размеры позволяют воздействовать на них с помощью специальных электродов без разрушения клетки.
Эти нейроны имеют развитую систему дендритов, которая распространяется по всей колонке, а так же аксон который в некоторых случаях является источником входящего или исходящего сигнала. Несмотря на то, что таких нейронов в первых слоях кортикальной колонки может быть несколько, все они работают как один нейроэлемент. Активность одной пирамидальной клетки зачастую означает активность всей группы клеток.
Эволюционное развитие нервной системы было ориентировано прежде на надежность и дублирование или распределение одного функционала на группы клеток является явлением повсеместным. Невозможно представить схему созданную природой, в которой гибель всего одной клетки приводило бы к нарушению работы целой функциональной единицы. Можно сказать, что активность группы пирамидальных клеток в колонке указывает на активность самой колонки.
Колонки можно разделить на два типа: формирующие сигнал и принимающие сигнал. Тип колонки зависит от того какие имеются аксоны у пирамидальных нейронов: афферентные или эфферентные. Если имеется афферентные аксоны, то есть приносящие сигнал, то колонка будет активироваться при получении сигнала, возможно ретрансляция сигнала, если будет и эфферентный аксон. При наличии эфферентных аксонов результатом активности колонки будет формирование исходящего сигнала.
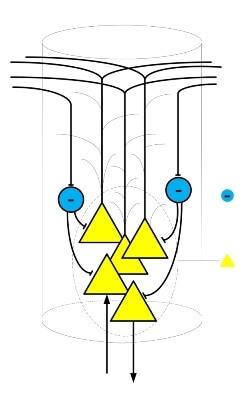
Дендриты пирамидальных нейронов тянутся к верхним слоям коры, где выходят за пределы своей кортикальной колонки, таким образом, осуществляется локальное взаимодействие между соседними колонками. Одна из форм локального взаимодействия это латеральное (боковое) торможение.
Торможение соседних колонок осуществляется посредством специальных тормозящих нейронов, которые входят в состав кортикальных колонок. Главным образом тормозящие нейроны оказывают своё действие на пирамидальные нейроны, препятствуя их активации.
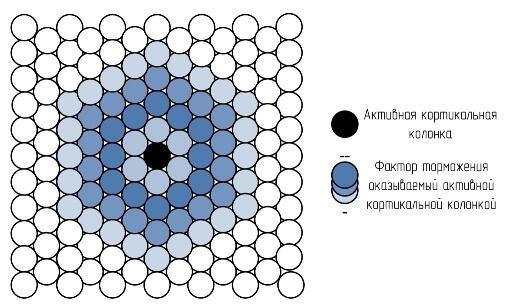
Латеральное торможение оказывается на соседние окружающие колонки. Оно позволяет сделать границы областей активных колонок более четкими и области активности более локализованными. Благодаря латеральному торможению возникает препятствие к сильному распространению возбуждения.
Помимо латерального торможения присутствует и латеральное побуждение. Настраивая баланс, между этими двумя факторами существует возможность очень тонко регулировать уровень общей активности в мозге. К примеру, для сна требуется снизить уровень активности для этого необходимо усилить латеральное торможение и ослабить побуждение. Это делается посредством специальных химических сигналов и медиаторов.
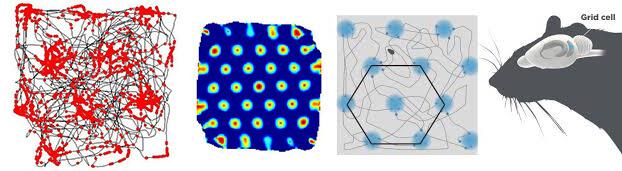
Не всегда действие на соседние колонки имеет форму равномерного концентричного распределения для энторинальной коры головного мозга свойственно распределение, которое дает возможность возбуждению легче распространяться в некой треугольной сетке. Это так называемые grid cell, которые помогают животными ориентироваться в пространстве, путем дополнительной модификации образа места пребывания.
Следующим элементом кортикальной колонки является слой из множества относительно маленьких звездчатых нейронов. Такой слой обычно называют зернистым. Нейроны этого слоя из-за своих размеров менее изучены, чем пирамидальные нейроны.
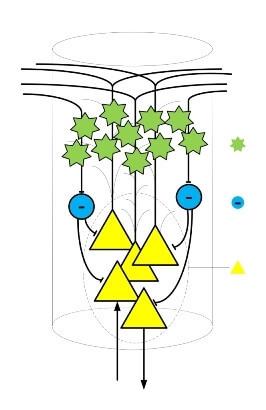
Именно нейронам этого слоя можно присвоить главную роль в вычислительных процессах мозга, благодаря их работе и происходит формирование ассоциативных связей и формирование образов. «Вычисления» происходят по принципам взаимопритяжения возбуждения. Нейроны зернистого слоя – это ассоциативный нейроэлемент.
А пирамидальным клеткам отводится роль простого сумматора, эти элементы как бы сепарируют, выделяют образы. Пирамидальные нейроны будут активироваться при возбуждении значительной части нейронов зернистого слоя колонки.
Связи между звездчатыми нейронами могут свободно выходить за границы своих кортикальных колонок, можно сказать, что зернистый слой является практически сплошным.
Память, консолидация памяти и бабушкины нейроны
Такой вариант кортикальной колонки и структуры коры очень прост и может показаться даже примитивным, но при использовании его в больших масштабах и при правильной настройке можно получить структуру с высокими вычислительными показателями. Природа всегда выбирала наиболее простые, надежные и эффективные решения и наша нервная система это не исключение их этого правила.
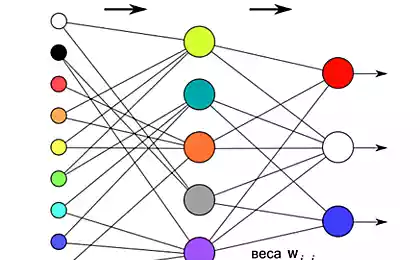
Зачастую сталкиваюсь с мнением, что нейрон подобен некоторому супервычислителю или квантовому компьютеру, который делает сложные вычисления, используя некие колебания ионов на своей мембране или квантовые механизмы. Даже алгоритм суммации в нейронах обычного персептрона является более сложным, чем в биологическом аналоге, в персептроне имеет значение от какого из синапсов приходят сигналы, а в биологическом важна лишь общая сумма воздействия. Стереотипное мышление о недосягаемой сложности мозга может препятствовать пониманию природы биологической системы.
Удивительно, но описанная структура является универсальной для различных типов областей коры: сенсорной, моторной и ассоциативной. Возможны вариации относительной толщины различных слоев в зависимости от функции области коры. К примеру, у моторной коры увеличен слой пирамидальных нейронов по отношению к зернистому слою, ведь сигналы моторной коры должны быть четкими и сильными. А для ассоциативных областей свойственно увеличенный слой звездчатых нейронов, чтобы обеспечить наибольшую гибкость в формировании ассоциативных рефлекторных дуг.
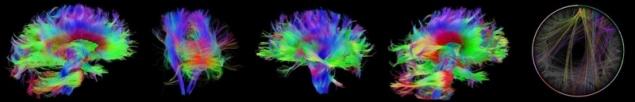
Области коры переплетены множеством связей, это осуществляется благодаря аксонам, длинным отросткам нейронов. Пучки аксонов формируют, нервны, которые является так называемым белым веществом. Эти нервы могут связывать как соседние области, так и области противоположных половин мозга. Причем архитектура этих связей обусловлена эволюционным развитием мозга и частично приобретенным опытом и наущением, но для разных людей картина этих связей будет сходна. Существует несколько научных проектов связанных с картированием этих связей, к примеру, Human Connectom Project.
Давайте рассмотрим то, по каким принципам организованны данные связи.
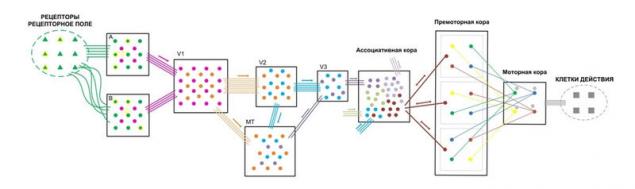
Представленная схема является лишь примером для понимания принципов организации. Реальные схемы в биологической нервной системе многократно сложнее.
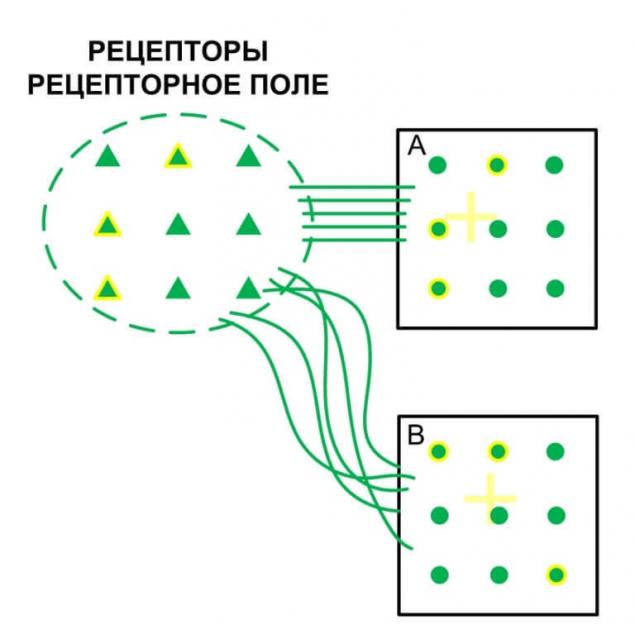
Представим некоторое рецепторное поле с рядом однотипных рецепторов, позволяющих получить об окружающей среде некую информацию. Определенные рецепторные поля формируют комбинированные сигналы, к примеру, сетчатка глаза. Такие сигналы требуют определенных возможностей анализа.
Представители-колонки данных рецепторов будут распределены по сенсорной коре с определенной плотностью с сохранением топологии расположения рецепторов в рецепторном поле (A). По принципам взаимного притяжения возбуждения на коре будут образовываться определенные участки возбуждения, что будет являться образом поступившего комбинированного сигнала.
Первичная сенсорная кора обычно имеет максимально высокую нейропластичность, т.е. любая комбинация возбужденных колонок будет обрабатываться без учёта ранее полученной информации. Полученный образ будет считываться другими колонками, также с определённой плотностью распределенными по сенсорной коре. Эти колонки будут передавать информации для дальнейшей обработки следующим областям коры.
Характер и плотность «считывающих» колонок дают определённый фильтр для полученных образов. Не сложно понять, что такой способ обработки приводит к значительной потере информации полученной от рецепторов, полученные образы не дают однозначной информации какие именно рецепторы были активированы. Эволюцией выбрано два пути устранения данной проблемы.
Во-первых, это избыточное количество рецепторов, что компенсируют падение информативности после обработки. Во-вторых, если продублировать информацию с рецепторов в другой области, но с нарушением топологии расположения колонок-представителей по отношению к расположению рецепторов в рецепторном поле (B), т.е. спутаем их.
То при множестве различных комбинаций активности рецепторов на двух областях будут формироваться различные комбинации образов, что дает больше информативности и больше признаков, которые мы можем выделить. Естественно следует понимать, что «спутывание» сигналов происходит строго определенным образом, к примеру, если «спутать» все поле сетчатки глаза никакой пользы это не даст.
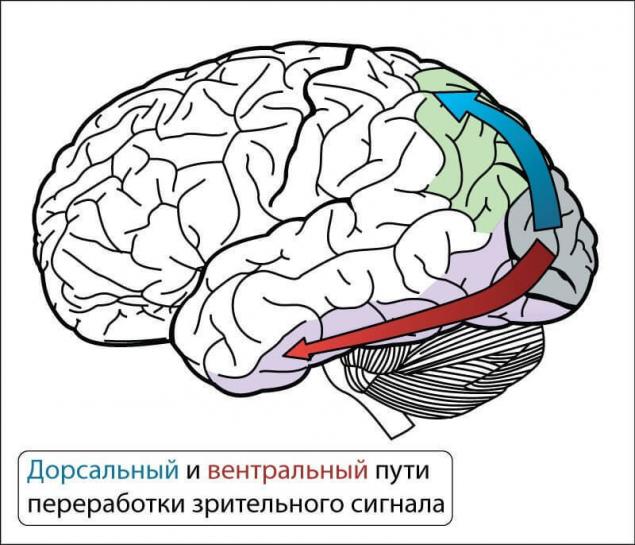
Спутывание происходит небольшими фрагментами рецепторных полей. И конечно мы можем, если это потребуется продублировать и спутать сигналы не один раз. В нервной системе примером этого разделения является дорсальный и вентральный пути переработки зрительного сигнала.
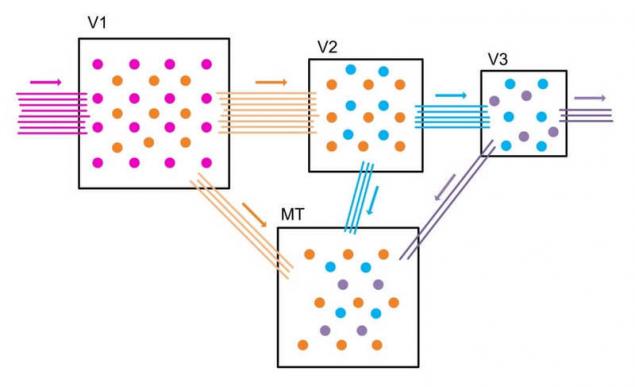
Общий принцип обработки информации корой больших полушарий это последовательная передача информации от области к области с понижением плотности связей. Причем с каждым последующим уровнем нейропластичность областей снижается, что добавляет к вычислениям память и опыт обработки предшествующей информации.
Тем самым из обрабатываемой информации могут выделяться некоторые ключевые признаки, которые будут связанны с конкретными нервными клетками. Проще говоря, на областях коры с более высоким уровнем обработки будет формироваться та форма распределения возбуждения, которая соответствует наиболее частой форме обрабатываемого сигнала.
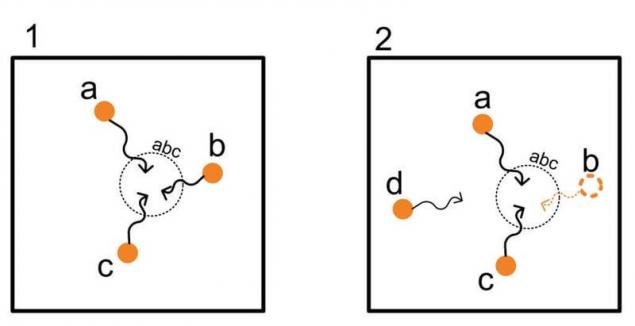
Сформированный образ «abc» комбинацией колонок a, b и c частым повторением или эмоциональным подкреплением, будет выполняться в дальнейшем даже при условии неполноты и зашумлённости активируемой комбинации.
На каждом уровне обработки информации затрачивается определенное время, если необходимо проанализировать динамику изменения информации во времени, то возможно продублирование части информации с каждого уровня на одну область.
Аналогом в биологии является область обработки зрительной информации MT (V5) средне-височная кора в которой собирается информация с областей V1, V2, V3…, эта область ответственна за восприятие движения. При повреждении этой области возникает акинетопсия – неспособность воспринимать движение.
Ассоциативные области устроены относительно просто, здесь располагаются представительства различных сенсорных и моторных областей. Причем для моторных представительств важно наличие двусторонних связей, либо это аксоны которые могут работать в оба направления, либо это соседствующие и совместно работающие колонки на обоих концах, либо два аксона одной колонки, работающие в различных направлениях.
Таких областей должно быть много с различным характером расположения представительств, чтобы возможность формирования рефлекторных дуг для различных комбинаций было равновозможным. На подобных областях формируются условные рефлексы, поэтому данные области должны обладать пониженной пластичностью.
Все связи в моторных и примоторных областях должны иметь двусторонний характер, это необходимо для формирования ассоциативных связей. По существу примоторная область должна формировать рефлекторные дуги последовательностей, от одного очага возбуждения к другому. Что бы было больше вариативности необходимо представительства, приводящие к действиям неоднократно перемешать, это похоже на ситуацию с сенсорной обработкой только в обратном порядке. Также все моторные действия взаимодействуют со специально устроенной областью координирующей действия во времени – мозжечком.
Представленные схемы являются сильным упрощением того как это утроено в мозге, и создание логических структур подобных нервной системе человека невозможно без участия специалистов в области нейробиологии и учёных изучающих коннектом.
А как же остальные слои?! – Действительно, я рассказал только про три слоя коры, но в мозге человека насчитывается шесть слоев в коре больших полушарий. Кора мозга оказался довольно успешным продуктом эволюции даже при наличии небольшого числа слоев. Принцип эволюции: то, что работает – не трогай.
Поэтому все новые слой в коре это надстройка к уже имеющимся слоям. Если разобраться в слоях мозга человека, то можно увидеть, что мы имеем не шесть видимых слоев, а два логических слоя, структура которых сходна и повторяется. Эволюция просто повторила уже имеющиеся структуру с целью увеличения производительности.
Пирамидальные клетки наружного слоя более мелкие, чем пирамидальные клетки первого слоя, а значит, в основном имеют более высокий порог чувствительности к активирующим факторам. Зернистые слоя будут работать в сходных условиях, но предположительно звездчатые нейроны наружного слоя имею более низкую пластичность, а значит, при некоторых условиях картины активности в зернистых слоях могут различаться, несмотря на то, что поступившие сигналы будут одинаковыми.
Благодаря этим двум логическим слоям возникает два режима активности кортикальной колонки. Первый: режим полной активности, кода пирамидальные клетки обоих слоев активируются, активируется вся колонка целиком. Второй: режим частичной активности, когда активируются только верхний дополнительный слой. Эти два режима работы колонки можно сравнить со способностью человека говорить в полный голос и шепотом, шепот – это частичная активность, а полный голос – это полная активность.
Что это дает? Для сенсорной коры – это дополнительный уровень обработки информации, а так же возможность работать с образами этих областей без активации через рецепторы. Другими словами дает возможность работать с фантазией. Для ассоциативных областей это дополнительный уровень абстракции, формирование ассоциаций между образами имеющих меньше общих признаков, так как порог чувствительности пирамидальных клеток дополнительного слоя более высок.
Для моторной и примоторной коры – это возможность проработать некоторые движения без непосредственного их выполнения. Только при полной активации колонки происходят действия, действия при частичной активации остаются в нашем воображении.
Конечно, есть области в мозге, которые управляют, работай режимов колонок, подобно тому, как мы можем очень легко сменить характер нашего говорения с шепота на полную силу. Если увеличить уровень торможения в колонке, то возможно, что она будет активироваться только частично, если наоборот растормозить колонку, то некоторые мысли могут сразу воплощаться в действия.
Фантазия и высокий уровень абстрактного мышления сделали человека самым успешным видом на Земле.
Даже если мы правильно настроим области и связи между ними, этого будет не достаточно для получения действующей модели. Необходимы безусловные рефлексы. Человек рождается с богатым набором рефлекторных механизмов старательно отобранных эволюцией.
Настройка безусловных рефлексов для модели является важным моментом, учитывая следующий факт, что обучение новым рефлексам происходит всегда на основе уже имеющихся рефлексов. Если какое-либо действие не будет вовлечено в какой-нибудь безусловный рефлекс, то обучится управлять этим действием, будет невозможно.
В биологических системах изначально не закладывается «четких» рефлексов. После рождения мы не можем точно управлять нашими конечностями или, к примеру, ходить. Это связанно с тем, что невозможно определить заранее некоторые параметры тела, размеры конечностей, их вес, усилие создаваемое мышцами и т.д. причем эти параметры еще и динамически изменяются в процессе роста организма.
Поэтому многие безусловные рефлексы имеют в ответе некоторое поле действий, а в заголовке поле активирующих его рецепторов. Так же закладывается эмоциональный механизм, связанный с безусловными рефлексами, который будет запускать рефлексы для корректировки в определённый момент развития.
Рассмотрим механизм корректировки рефлексов на примере детского лепета. В соответствии с определенным этапом развития запускается механизм лепетания, т.е. происходит практически спонтанный запуск «нечетких» рефлексов. При их запуске ребёнок начинает произносить различные звуки, иногда эти же рефлексы провоцируются и услышанным звуком со стороны.
Произнесенные звуки зачастую не соответствуют ожидаемым, т.е. не соответствуют звукам запускающим рефлекс, или запускающим гипотетически. Ребёнок слышит издаваемые им самим звуки, получая обратную связь между командой и результирующим действием. Далее в силу вступает эмоциональный механизм новизны, который связывает центр потребности в новизне с речевым актом, который дает новую ассоциацию между слышимым звуком и внутренним мотивом, побудившим к действию.
Что приводит к многократному повторению действия приводящего к насыщению чувства новизны. Утверждается, что ребёнок в период лепета произносит все звуки всех языков на Земле. Многократное повторение звуков приводит к формированию чётких образов действий в соответствии с желанным результатом.
Аналогично происходит и овладение моторно-двигательной системой. Изначально движения младенцев практически хаотичны, присутствует только увеличение двигательной активности как реакция на эмоциональный стимул. Но со временем происходит сопоставление движений и зрительного восприятия, тактильного и восприятия положения тела.
Некоторые безусловные рефлексы не так примитивны, в определённых случаях в нервной системе заложены шаблонные образы, и перенести такие шаблоны из биологических систем в компьютерную модель практически невозможно. Человек имеет врожденную способность к распознаванию эмоций и движений особей своего вида. Поэтому в отношении некоторых аспектов обучения будет необходимость применять некоторые обходные пути.
Что бы миновать процесс длительного обучения моторно-двигательной системы через многочисленные попытки ползания, вставаний, ходьбы и череду падений для роботов андройдов возможно применить метод перехвата управления.

Человек может передать свой опыт управления телом роботу посредством специальных устройств и технологий перехвата движений. В модели нервной системы робота с перехваченным управлением при моторных действиях будут активироваться соответствующие представительства, так если бы эти движения выполнял сам робот.
Благодаря чему формировались бы необходимые образы и ассоциативные связи. К примеру, во время обучения при команде: «Подыми руки» — обучающий в режиме перехвата движений сам подымает свои руки, то это приводило бы формированию условного рефлекса между командой и действием, а также формировалась бы ассоциативная связь между командой и образом сформированным обработкой датчиков положения тела.
ПОДПИСЫВАЙТЕСЬ на НАШ youtube канал, что позволяет смотреть онлайн, скачать с ютуб бесплатно видео об оздоровлении, омоложении человека. Любовь к окружающим и к себе, как чувство высоких вибраций — важный фактор
В процессе обучения электронной модели мозга всегда возможно управлять пластичностью желаемых областей, а так же есть возможность заглянуть «внутрь» процесса обучения, и выделять, обозначать и укреплять получаемые образы. Что должно существенно ускорить процесс обучения искусственных нервных систем в соотношении с обучением человека. Как уже стало ясно, модель, построенная по описанным принципам, будет обучаться по большей части подобно человеку, без разделения обучения и взаимодействия с окружающей средой.опубликовано
Ставьте ЛАЙКИ, делитесь с ДРУЗЬЯМИ!
www.youtube.com/channel/UCXd71u0w04qcwk32c8kY2BA/videos
Подпишитесь — www.facebook.com//
Источник: geektimes.ru/post/280604/
Недостаточно взять некую массу связанных нейронов (даже сложив слоями) подключить к ней датчики и выводы и получить хоть какое-то подобие мозга. Главным полем работы эволюции на протяжении миллионов лет является не нейрон, а структура и внутренняя организация нервных клеток в нервной системе.

Давайте разберемся в том, как организована кора больших полушарий, структура благодаря которой Человек стал самым успешным видом на Земле.
Известно, что кора больших полушарий мозга человека состоит из шести условных слоев, но к такой форме она пришла за многие этапы эволюции. Поэтому для начала мы будем рассматривать упрощённый вариант с 2 – 3 слоями, ведь и такой вариант в природе имеет место. Любую биологическую структуру выгоднее изучать с позиции её эволюционного развития. Эволюция шла двумя путями развития коры: это увеличение количества слоёв и увеличение общей площади коры.

Второй особенностью коры является наличие так называемых кортикальных колонок. Можно сказать, что кортикальная колонка это логическая единица структуры коры. Рост самой коры во время эмбрионального развития осуществляется делением, самокопированием целых колонок. И это логично, если мы хотим сохранить некую структуру при росте.

В основе кортикальных колонок лежат пирамидальные нейроны. Пирамидальные нейроны одни из самых крупных нейронов нервной системы, поэтому наиболее изученные, так как их размеры позволяют воздействовать на них с помощью специальных электродов без разрушения клетки.
Эти нейроны имеют развитую систему дендритов, которая распространяется по всей колонке, а так же аксон который в некоторых случаях является источником входящего или исходящего сигнала. Несмотря на то, что таких нейронов в первых слоях кортикальной колонки может быть несколько, все они работают как один нейроэлемент. Активность одной пирамидальной клетки зачастую означает активность всей группы клеток.
Эволюционное развитие нервной системы было ориентировано прежде на надежность и дублирование или распределение одного функционала на группы клеток является явлением повсеместным. Невозможно представить схему созданную природой, в которой гибель всего одной клетки приводило бы к нарушению работы целой функциональной единицы. Можно сказать, что активность группы пирамидальных клеток в колонке указывает на активность самой колонки.
Колонки можно разделить на два типа: формирующие сигнал и принимающие сигнал. Тип колонки зависит от того какие имеются аксоны у пирамидальных нейронов: афферентные или эфферентные. Если имеется афферентные аксоны, то есть приносящие сигнал, то колонка будет активироваться при получении сигнала, возможно ретрансляция сигнала, если будет и эфферентный аксон. При наличии эфферентных аксонов результатом активности колонки будет формирование исходящего сигнала.

Дендриты пирамидальных нейронов тянутся к верхним слоям коры, где выходят за пределы своей кортикальной колонки, таким образом, осуществляется локальное взаимодействие между соседними колонками. Одна из форм локального взаимодействия это латеральное (боковое) торможение.
Торможение соседних колонок осуществляется посредством специальных тормозящих нейронов, которые входят в состав кортикальных колонок. Главным образом тормозящие нейроны оказывают своё действие на пирамидальные нейроны, препятствуя их активации.
Латеральное торможение оказывается на соседние окружающие колонки. Оно позволяет сделать границы областей активных колонок более четкими и области активности более локализованными. Благодаря латеральному торможению возникает препятствие к сильному распространению возбуждения.
Помимо латерального торможения присутствует и латеральное побуждение. Настраивая баланс, между этими двумя факторами существует возможность очень тонко регулировать уровень общей активности в мозге. К примеру, для сна требуется снизить уровень активности для этого необходимо усилить латеральное торможение и ослабить побуждение. Это делается посредством специальных химических сигналов и медиаторов.

Не всегда действие на соседние колонки имеет форму равномерного концентричного распределения для энторинальной коры головного мозга свойственно распределение, которое дает возможность возбуждению легче распространяться в некой треугольной сетке. Это так называемые grid cell, которые помогают животными ориентироваться в пространстве, путем дополнительной модификации образа места пребывания.

Следующим элементом кортикальной колонки является слой из множества относительно маленьких звездчатых нейронов. Такой слой обычно называют зернистым. Нейроны этого слоя из-за своих размеров менее изучены, чем пирамидальные нейроны.

Именно нейронам этого слоя можно присвоить главную роль в вычислительных процессах мозга, благодаря их работе и происходит формирование ассоциативных связей и формирование образов. «Вычисления» происходят по принципам взаимопритяжения возбуждения. Нейроны зернистого слоя – это ассоциативный нейроэлемент.
А пирамидальным клеткам отводится роль простого сумматора, эти элементы как бы сепарируют, выделяют образы. Пирамидальные нейроны будут активироваться при возбуждении значительной части нейронов зернистого слоя колонки.
Связи между звездчатыми нейронами могут свободно выходить за границы своих кортикальных колонок, можно сказать, что зернистый слой является практически сплошным.
Память, консолидация памяти и бабушкины нейроны
Такой вариант кортикальной колонки и структуры коры очень прост и может показаться даже примитивным, но при использовании его в больших масштабах и при правильной настройке можно получить структуру с высокими вычислительными показателями. Природа всегда выбирала наиболее простые, надежные и эффективные решения и наша нервная система это не исключение их этого правила.
Зачастую сталкиваюсь с мнением, что нейрон подобен некоторому супервычислителю или квантовому компьютеру, который делает сложные вычисления, используя некие колебания ионов на своей мембране или квантовые механизмы. Даже алгоритм суммации в нейронах обычного персептрона является более сложным, чем в биологическом аналоге, в персептроне имеет значение от какого из синапсов приходят сигналы, а в биологическом важна лишь общая сумма воздействия. Стереотипное мышление о недосягаемой сложности мозга может препятствовать пониманию природы биологической системы.
Удивительно, но описанная структура является универсальной для различных типов областей коры: сенсорной, моторной и ассоциативной. Возможны вариации относительной толщины различных слоев в зависимости от функции области коры. К примеру, у моторной коры увеличен слой пирамидальных нейронов по отношению к зернистому слою, ведь сигналы моторной коры должны быть четкими и сильными. А для ассоциативных областей свойственно увеличенный слой звездчатых нейронов, чтобы обеспечить наибольшую гибкость в формировании ассоциативных рефлекторных дуг.
Области коры переплетены множеством связей, это осуществляется благодаря аксонам, длинным отросткам нейронов. Пучки аксонов формируют, нервны, которые является так называемым белым веществом. Эти нервы могут связывать как соседние области, так и области противоположных половин мозга. Причем архитектура этих связей обусловлена эволюционным развитием мозга и частично приобретенным опытом и наущением, но для разных людей картина этих связей будет сходна. Существует несколько научных проектов связанных с картированием этих связей, к примеру, Human Connectom Project.

Давайте рассмотрим то, по каким принципам организованны данные связи.

Представленная схема является лишь примером для понимания принципов организации. Реальные схемы в биологической нервной системе многократно сложнее.

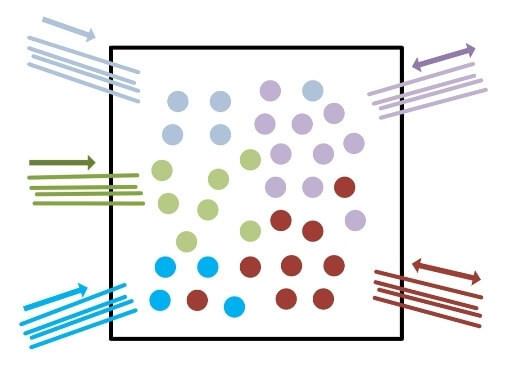
Представим некоторое рецепторное поле с рядом однотипных рецепторов, позволяющих получить об окружающей среде некую информацию. Определенные рецепторные поля формируют комбинированные сигналы, к примеру, сетчатка глаза. Такие сигналы требуют определенных возможностей анализа.
Представители-колонки данных рецепторов будут распределены по сенсорной коре с определенной плотностью с сохранением топологии расположения рецепторов в рецепторном поле (A). По принципам взаимного притяжения возбуждения на коре будут образовываться определенные участки возбуждения, что будет являться образом поступившего комбинированного сигнала.
Первичная сенсорная кора обычно имеет максимально высокую нейропластичность, т.е. любая комбинация возбужденных колонок будет обрабатываться без учёта ранее полученной информации. Полученный образ будет считываться другими колонками, также с определённой плотностью распределенными по сенсорной коре. Эти колонки будут передавать информации для дальнейшей обработки следующим областям коры.
Характер и плотность «считывающих» колонок дают определённый фильтр для полученных образов. Не сложно понять, что такой способ обработки приводит к значительной потере информации полученной от рецепторов, полученные образы не дают однозначной информации какие именно рецепторы были активированы. Эволюцией выбрано два пути устранения данной проблемы.
Во-первых, это избыточное количество рецепторов, что компенсируют падение информативности после обработки. Во-вторых, если продублировать информацию с рецепторов в другой области, но с нарушением топологии расположения колонок-представителей по отношению к расположению рецепторов в рецепторном поле (B), т.е. спутаем их.
То при множестве различных комбинаций активности рецепторов на двух областях будут формироваться различные комбинации образов, что дает больше информативности и больше признаков, которые мы можем выделить. Естественно следует понимать, что «спутывание» сигналов происходит строго определенным образом, к примеру, если «спутать» все поле сетчатки глаза никакой пользы это не даст.
Спутывание происходит небольшими фрагментами рецепторных полей. И конечно мы можем, если это потребуется продублировать и спутать сигналы не один раз. В нервной системе примером этого разделения является дорсальный и вентральный пути переработки зрительного сигнала.


Общий принцип обработки информации корой больших полушарий это последовательная передача информации от области к области с понижением плотности связей. Причем с каждым последующим уровнем нейропластичность областей снижается, что добавляет к вычислениям память и опыт обработки предшествующей информации.
Тем самым из обрабатываемой информации могут выделяться некоторые ключевые признаки, которые будут связанны с конкретными нервными клетками. Проще говоря, на областях коры с более высоким уровнем обработки будет формироваться та форма распределения возбуждения, которая соответствует наиболее частой форме обрабатываемого сигнала.

Сформированный образ «abc» комбинацией колонок a, b и c частым повторением или эмоциональным подкреплением, будет выполняться в дальнейшем даже при условии неполноты и зашумлённости активируемой комбинации.
На каждом уровне обработки информации затрачивается определенное время, если необходимо проанализировать динамику изменения информации во времени, то возможно продублирование части информации с каждого уровня на одну область.
Аналогом в биологии является область обработки зрительной информации MT (V5) средне-височная кора в которой собирается информация с областей V1, V2, V3…, эта область ответственна за восприятие движения. При повреждении этой области возникает акинетопсия – неспособность воспринимать движение.

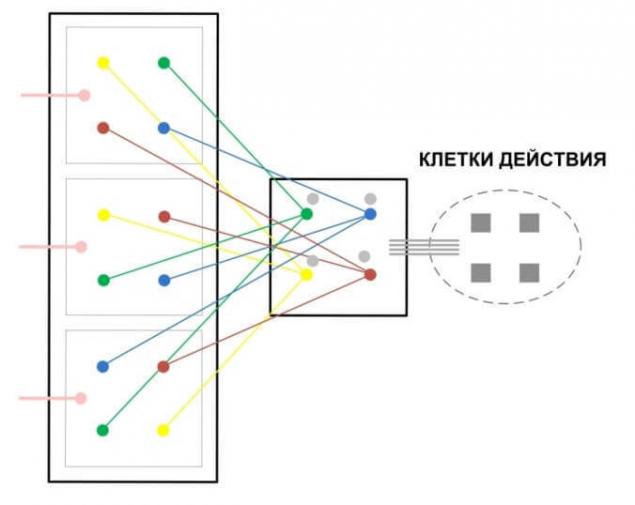
Ассоциативные области устроены относительно просто, здесь располагаются представительства различных сенсорных и моторных областей. Причем для моторных представительств важно наличие двусторонних связей, либо это аксоны которые могут работать в оба направления, либо это соседствующие и совместно работающие колонки на обоих концах, либо два аксона одной колонки, работающие в различных направлениях.
Таких областей должно быть много с различным характером расположения представительств, чтобы возможность формирования рефлекторных дуг для различных комбинаций было равновозможным. На подобных областях формируются условные рефлексы, поэтому данные области должны обладать пониженной пластичностью.

Все связи в моторных и примоторных областях должны иметь двусторонний характер, это необходимо для формирования ассоциативных связей. По существу примоторная область должна формировать рефлекторные дуги последовательностей, от одного очага возбуждения к другому. Что бы было больше вариативности необходимо представительства, приводящие к действиям неоднократно перемешать, это похоже на ситуацию с сенсорной обработкой только в обратном порядке. Также все моторные действия взаимодействуют со специально устроенной областью координирующей действия во времени – мозжечком.
Представленные схемы являются сильным упрощением того как это утроено в мозге, и создание логических структур подобных нервной системе человека невозможно без участия специалистов в области нейробиологии и учёных изучающих коннектом.
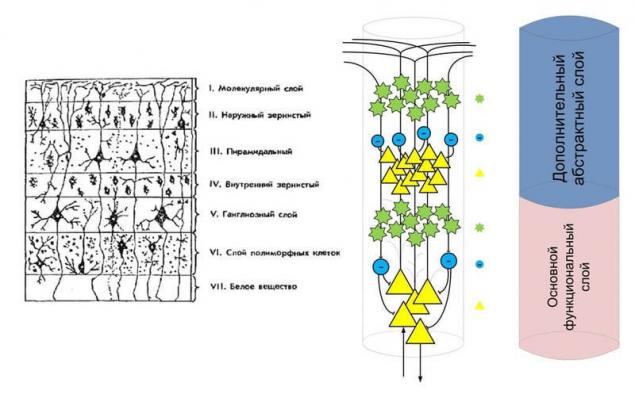
А как же остальные слои?! – Действительно, я рассказал только про три слоя коры, но в мозге человека насчитывается шесть слоев в коре больших полушарий. Кора мозга оказался довольно успешным продуктом эволюции даже при наличии небольшого числа слоев. Принцип эволюции: то, что работает – не трогай.
Поэтому все новые слой в коре это надстройка к уже имеющимся слоям. Если разобраться в слоях мозга человека, то можно увидеть, что мы имеем не шесть видимых слоев, а два логических слоя, структура которых сходна и повторяется. Эволюция просто повторила уже имеющиеся структуру с целью увеличения производительности.

Пирамидальные клетки наружного слоя более мелкие, чем пирамидальные клетки первого слоя, а значит, в основном имеют более высокий порог чувствительности к активирующим факторам. Зернистые слоя будут работать в сходных условиях, но предположительно звездчатые нейроны наружного слоя имею более низкую пластичность, а значит, при некоторых условиях картины активности в зернистых слоях могут различаться, несмотря на то, что поступившие сигналы будут одинаковыми.
Благодаря этим двум логическим слоям возникает два режима активности кортикальной колонки. Первый: режим полной активности, кода пирамидальные клетки обоих слоев активируются, активируется вся колонка целиком. Второй: режим частичной активности, когда активируются только верхний дополнительный слой. Эти два режима работы колонки можно сравнить со способностью человека говорить в полный голос и шепотом, шепот – это частичная активность, а полный голос – это полная активность.
Что это дает? Для сенсорной коры – это дополнительный уровень обработки информации, а так же возможность работать с образами этих областей без активации через рецепторы. Другими словами дает возможность работать с фантазией. Для ассоциативных областей это дополнительный уровень абстракции, формирование ассоциаций между образами имеющих меньше общих признаков, так как порог чувствительности пирамидальных клеток дополнительного слоя более высок.
Для моторной и примоторной коры – это возможность проработать некоторые движения без непосредственного их выполнения. Только при полной активации колонки происходят действия, действия при частичной активации остаются в нашем воображении.
Конечно, есть области в мозге, которые управляют, работай режимов колонок, подобно тому, как мы можем очень легко сменить характер нашего говорения с шепота на полную силу. Если увеличить уровень торможения в колонке, то возможно, что она будет активироваться только частично, если наоборот растормозить колонку, то некоторые мысли могут сразу воплощаться в действия.
Фантазия и высокий уровень абстрактного мышления сделали человека самым успешным видом на Земле.
Даже если мы правильно настроим области и связи между ними, этого будет не достаточно для получения действующей модели. Необходимы безусловные рефлексы. Человек рождается с богатым набором рефлекторных механизмов старательно отобранных эволюцией.
Настройка безусловных рефлексов для модели является важным моментом, учитывая следующий факт, что обучение новым рефлексам происходит всегда на основе уже имеющихся рефлексов. Если какое-либо действие не будет вовлечено в какой-нибудь безусловный рефлекс, то обучится управлять этим действием, будет невозможно.
В биологических системах изначально не закладывается «четких» рефлексов. После рождения мы не можем точно управлять нашими конечностями или, к примеру, ходить. Это связанно с тем, что невозможно определить заранее некоторые параметры тела, размеры конечностей, их вес, усилие создаваемое мышцами и т.д. причем эти параметры еще и динамически изменяются в процессе роста организма.
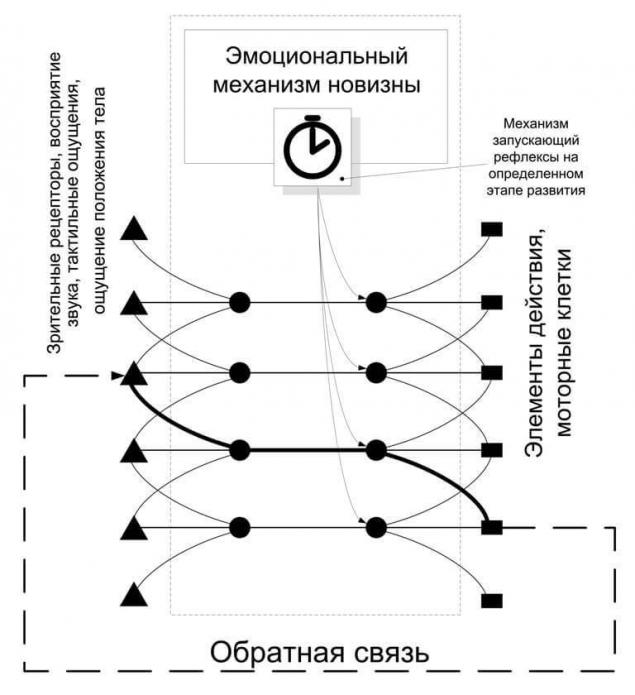
Поэтому многие безусловные рефлексы имеют в ответе некоторое поле действий, а в заголовке поле активирующих его рецепторов. Так же закладывается эмоциональный механизм, связанный с безусловными рефлексами, который будет запускать рефлексы для корректировки в определённый момент развития.

Рассмотрим механизм корректировки рефлексов на примере детского лепета. В соответствии с определенным этапом развития запускается механизм лепетания, т.е. происходит практически спонтанный запуск «нечетких» рефлексов. При их запуске ребёнок начинает произносить различные звуки, иногда эти же рефлексы провоцируются и услышанным звуком со стороны.
Произнесенные звуки зачастую не соответствуют ожидаемым, т.е. не соответствуют звукам запускающим рефлекс, или запускающим гипотетически. Ребёнок слышит издаваемые им самим звуки, получая обратную связь между командой и результирующим действием. Далее в силу вступает эмоциональный механизм новизны, который связывает центр потребности в новизне с речевым актом, который дает новую ассоциацию между слышимым звуком и внутренним мотивом, побудившим к действию.
Что приводит к многократному повторению действия приводящего к насыщению чувства новизны. Утверждается, что ребёнок в период лепета произносит все звуки всех языков на Земле. Многократное повторение звуков приводит к формированию чётких образов действий в соответствии с желанным результатом.
Аналогично происходит и овладение моторно-двигательной системой. Изначально движения младенцев практически хаотичны, присутствует только увеличение двигательной активности как реакция на эмоциональный стимул. Но со временем происходит сопоставление движений и зрительного восприятия, тактильного и восприятия положения тела.
Некоторые безусловные рефлексы не так примитивны, в определённых случаях в нервной системе заложены шаблонные образы, и перенести такие шаблоны из биологических систем в компьютерную модель практически невозможно. Человек имеет врожденную способность к распознаванию эмоций и движений особей своего вида. Поэтому в отношении некоторых аспектов обучения будет необходимость применять некоторые обходные пути.
Что бы миновать процесс длительного обучения моторно-двигательной системы через многочисленные попытки ползания, вставаний, ходьбы и череду падений для роботов андройдов возможно применить метод перехвата управления.

Человек может передать свой опыт управления телом роботу посредством специальных устройств и технологий перехвата движений. В модели нервной системы робота с перехваченным управлением при моторных действиях будут активироваться соответствующие представительства, так если бы эти движения выполнял сам робот.
Благодаря чему формировались бы необходимые образы и ассоциативные связи. К примеру, во время обучения при команде: «Подыми руки» — обучающий в режиме перехвата движений сам подымает свои руки, то это приводило бы формированию условного рефлекса между командой и действием, а также формировалась бы ассоциативная связь между командой и образом сформированным обработкой датчиков положения тела.
ПОДПИСЫВАЙТЕСЬ на НАШ youtube канал, что позволяет смотреть онлайн, скачать с ютуб бесплатно видео об оздоровлении, омоложении человека. Любовь к окружающим и к себе, как чувство высоких вибраций — важный фактор
В процессе обучения электронной модели мозга всегда возможно управлять пластичностью желаемых областей, а так же есть возможность заглянуть «внутрь» процесса обучения, и выделять, обозначать и укреплять получаемые образы. Что должно существенно ускорить процесс обучения искусственных нервных систем в соотношении с обучением человека. Как уже стало ясно, модель, построенная по описанным принципам, будет обучаться по большей части подобно человеку, без разделения обучения и взаимодействия с окружающей средой.опубликовано
Как гены определяют наше поведение
«Кто мы есть на самом деле?»: лучшая подборка TED-лекций учёных и философов
Ставьте ЛАЙКИ, делитесь с ДРУЗЬЯМИ!
www.youtube.com/channel/UCXd71u0w04qcwk32c8kY2BA/videos
Подпишитесь — www.facebook.com//
Источник: geektimes.ru/post/280604/
Портал БАШНЯ. Копирование, Перепечатка возможна при указании активной ссылки на данную страницу.






























Остеохондроз: как ПРАВИЛЬНО сидеть, стоять, и поднимать тяжести
Письмо директора школы, которое стоит прочитать всем родителям