Trivia pensamiento o artículo sobre las espinas dendríticas
 Bashny.Net
Bashny.Net
Hace unos meses publicó una serie de artículos bajo el título «La lógica de pensar» . Con optimismo previsto continuar después de un mes o dos. Pero la vida ha hecho ajustes. Modelado modelo patrón de onda corteza dio resultados interesantes por lo que tuvimos que posponer todo lo demás, incluyendo la continuación del ciclo de escritura Habra.
Sin embargo, no hace mucho tiempo, escribí y publiqué en el artículo pre-impresión ( http://arxiv.org/abs/1406.6901 ). En cierto modo, puede ser interesante para aquellos que interesó previamente en el modelo de onda. Recordemos que el modelo de punto clave - una declaración de que las neuronas son capaces de memorizar y aprender no una sola imagen, describió sus pesos de las sinapsis, sino también una gran cantidad de otros de que las señales de imagen. Por supuesto, una complicación de una neurona como está en desacuerdo con muchas de las teorías existentes y requiere algo más que un estudio serio. A continuación, yo, simplemente, y trato de describir uno de los que figuran en los argumentos a favor de mi modelo.
Este artículo no debe ser tomado como una continuación del ciclo, es más bien una precuela a la misma. En cuanto a mí, he llamado a este argumento sobre el papel clave de las espinas dendríticas.
Comenzamos con la recurrencia conocido.
En el estado de reposo entre el interior y el exterior de la diferencia de potencial de neuronas - el potencial de membrana de alrededor de 70 mV. Está formada por las moléculas de proteínas que actúan como bombas de iones. Como resultado, la membrana asume la polarización para los que la carga negativa se acumula dentro de las células, y positivo exterior.
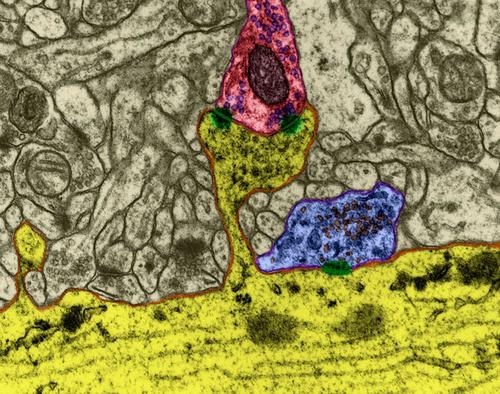
La superficie está cubierta con una neurona procesos de ramificación - dendritas. Para el cuerpo de la neurona y las dendritas son adyacentes a sus terminaciones axonales de otras neuronas. Sus articulaciones se denominan sinapsis. A través de la interacción de la neurona sináptica puede responder a las señales entrantes y bajo ciertas circunstancias para generar su propio impulso, llamado espiga.
La transmisión de señales en las sinapsis que ocurre a través de la liberación de neurotransmisores. Cuando un impulso nervioso a lo largo del axón entra en la terminal presináptica, que se libere de los neurotransmisores de molécula de vesículas sinápticas que son típicos de esta sinapsis. En la membrana de una neurona que recibe los receptores de señal colocado que interactúan con un neurotransmisor.
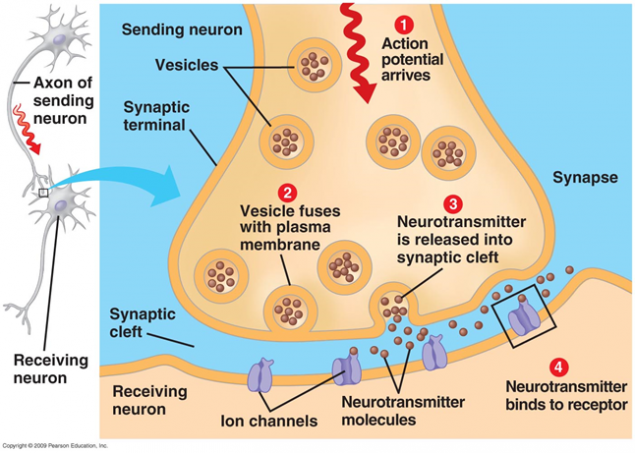
Figura 1. Sinapsis
Receptores localizados en la hendidura sináptica son en su mayoría ionotrópicos. Es decir, que son los mismos canales de iones, la capacidad de mover iones a través de la membrana de la neurona. Los neurotransmisores actúan como receptores que sus canales iónicos abiertos. Como resultado, una membrana se despolariza, o hiperpolarización - dependiendo de qué canales se ven afectados, y, en consecuencia, cualquier tipo de sinapsis. En las sinapsis excitadoras abre canales de cationes, principalmente permeables dentro de la célula - membrana se despolariza. En las sinapsis inhibitorias canales abiertos que dan cationes de las células, lo que conduce a la membrana hiperpolarización.
En ciertas circunstancias, las sinapsis pueden cambiar su sensibilidad se llama la plasticidad sináptica. Esto conduce al hecho de que algunas de las sinapsis se vuelven más y otras menos susceptibles a señales externas.
Al mismo tiempo, las sinapsis de las neuronas reciben muchas señales. Sinapsis inhibidoras cambian el potencial de membrana hacia la acumulación de carga dentro de la jaula. La activación de las sinapsis, por el contrario, tratar de calmar la neurona. Cuando el total excede el umbral de despolarización de la iniciación, una descarga, se llama un potencial de acción, o pico.
Después de la liberación de neurotransmisores mecanismos especiales de garantizar su reciclado y re-absorción, lo que conduce a la liquidación de brecha sináptica y el espacio que rodea la sinapsis. Durante el período refractario, viene después de un pico, la neurona no es capaz de generar nuevos impulsos. La duración de este período determina la frecuencia máxima de oscilación, que es capaz de una neurona.
Ahora describir los hechos menos conocidos.
Cuando un potencial de acción se propaga a lo largo del axón, la neurona alcanza el receptor, que provoca la liberación de neurotransmisores en la hendidura sináptica. Estos mediadores determinar la contribución de la sinapsis en el cambio total en el potencial de membrana de la neurona que recibe la señal. Pero parte de mediadores cae fuera de la brecha sináptica y llenando el espacio formado por las neuronas y células gliales circundantes. Este fenómeno se denomina derrame (spillover (Esp.) - Desbordamiento, desbordamiento) (Kullmann, 2000). Además, los mediadores se emiten terminales de los axones no sinápticos y células gliales (Figura 2). La concentración de sináptica de neurotransmisores exterior es mucho menor que en las hendiduras sinápticas. Sin embargo, es en estos neurotransmisores "rasteksheysya" masa de intereses ocultos.
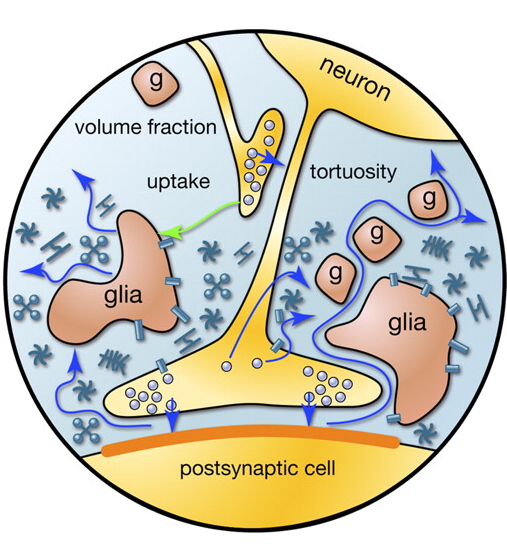
Figura 2. Fuentes de mediadores fuera de la hendidura sináptica (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
Vamos a tratar de estimar el número y la estructura de las fuentes de emisión de neurotransmisores fuera de las sinapsis. Para ello se utilizan las estimaciones cuantitativas de los parámetros de la corteza, en la siguiente tabla (Tabla 1) (Braitenberg V., Schuz A., 1998).
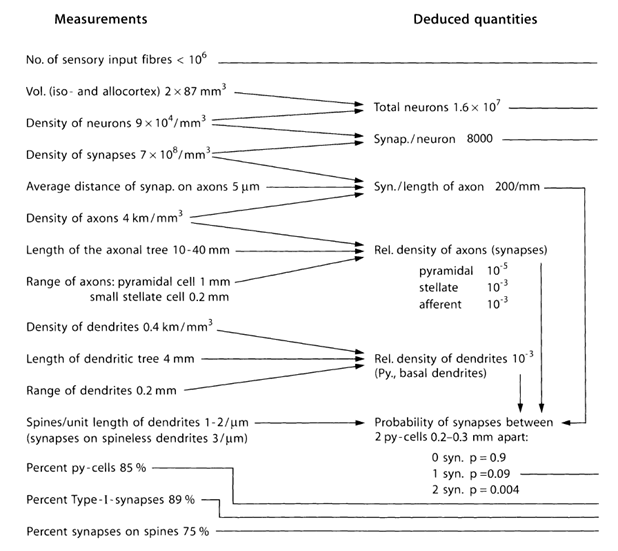
Tabla 1. Cuadro resumen de los parámetros obtenidos en el estudio de cerebro de ratón (células py - célula piramidal, de tipo I - las sinapsis entre las células piramidales) (Braitenberg V., Schuz A., 1998)
Permítanme que les recuerde que la mayoría de las sinapsis (90-95%) no está en el cuerpo de la neurona y sus dendritas. Las dendritas - a procesos de ramificación delgada que forma el llamado árbol dendrítico de la neurona. Las figuras de abajo árboles dendríticas resumen en negro, gris y axones. Para los diferentes tipos de neuronas forman árboles dendríticas son diferentes, pero el principio general se conserva: árbol dendrítico consiste en un conjunto de procesos de ramificación, con la mayor densidad de las conexiones sinápticas de una neurona cae en una pequeña región del espacio. Para los tipos básicos de neuronas, que es de aproximadamente 200 m (Figura 3, Figura 4).
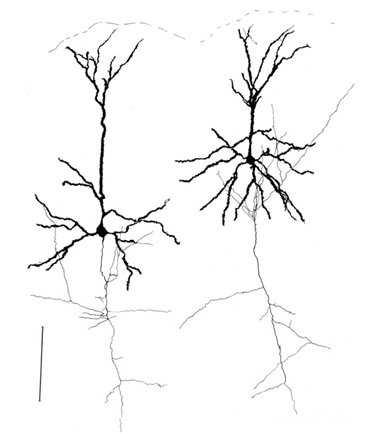
Figura 3. Estructura de la línea de las neuronas estrelladas - 0,1 mm (Braitenberg, 1978)
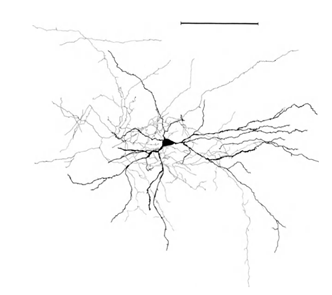
Figura 4. Estructura de las neuronas piramidales, regla - 0,1 mm (Braitenberg, 1978)
De ramificación de los axones de neuronas forman contactos (sinapsis) con las dendritas de otras neuronas. La distancia media entre las sinapsis en las dendritas - 0,5 micrómetros. La distancia media entre las sinapsis en los axones - 5 micrómetros, que es 10 veces mayor. No es sorprendente que los axones se obtienen aproximadamente 10 veces más que las dendritas.
La mayoría (75%) de las sinapsis localizados en las espinas dendríticas que es más característico de las células piramidales (Figura 5).
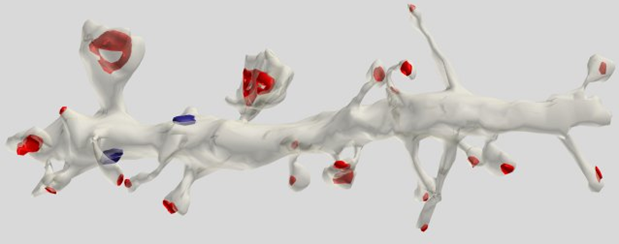
Figura 5. Segmento de dendritas de las células piramidales. Rojo marcó sinapsis en espinas, azul - en tronco dendríticas (Dr. Kristen M. Harris)
Las simulaciones por ordenador realizadas sobre la base de anatómica real y datos fisiológicos mostraron que, por ejemplo, el glutamato se puede extender más allá de la hendidura sináptica en cantidades suficientes para activar los receptores de NMDA dentro comparable con la distancia entre las sinapsis vecinas (0,5 micras) (Rusakov DA, Kullmann DM, 1998). Se puede suponer que una concentración significativa de los neurotransmisores después de derrame ocurre en el sitio de la longitud de las dendritas del orden de 1-2 micras. En este sitio puede ser localizado aproximadamente dos a cuatro sinapsis pertenecientes a la dendrita.
Si usted toma una porción de la longitud de las dendritas de 5 m (Figura 6), el número esperado de las sinapsis en él será de unos 10.
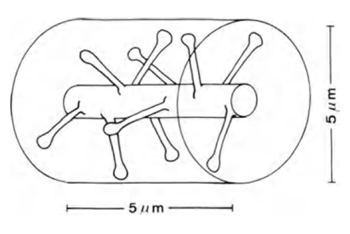
Figura 6. Parcela de dendritas (Braitenberg V., Schuz A., 1998)
Pero algunas ramas dendríticas de las neuronas están estrechamente entrelazadas con las ramas de otras neuronas. Probaron el uno al otro muy cerca. Debido a la altura de las espinas dendríticas sinapsis pertenecientes a la misma dendrita, puede estar más cerca de la superficie de la dendrita de otra que sus propios sinapsis.
Si las sinapsis se distribuyen de manera uniforme en el espacio de la corteza, el volumen cilíndrico de altura de 5 mm y un diámetro de 5 mm, también (foto de arriba) a una distribución de la densidad de las sinapsis 7x10 8 sup> / mm 3 sup> hit a alrededor de 100 sinapsis. Es decir, 10 veces más de lo que se encuentra directamente en la rama en sí. De hecho, una parte sustancial del volumen del cerebro ocupada por las células gliales y las neuronas del cuerpo, lo que aumenta aún más la densidad de empaquetamiento de las sinapsis. Sin embargo, las neuronas emplean diferentes neurotransmisores, que también debe ser considerado.
Ahora trata de entender el significado de una estructura de relaciones tales en términos de la distribución de la densidad extrasynaptic neurotransmisor. Para ello utilizamos un modelo simplificado. Tome condicional volumen neuronas circundantes y enumerar sus neuronas miembros. Cada una de estas neuronas sería:
• múltiples contactos sinápticos con las dendritas de las neuronas seleccionadas;
• Número de "ajuste", es decir, lugares en los que las sinapsis con otras neuronas están situadas en las proximidades de las dendritas de las neuronas seleccionadas.
Imagínese árbol dendrítico como una rama larga con las fuentes convencionales distribuidas uniformemente (Figura 7). Para cada fuente en este hilo, se puede especificar el número de neuronas del espacio circundante, es responsable de la misma. Cada una de las neuronas del medio ambiente tendrá múltiples contactos de fuente, distribuidos al azar a lo largo de las dendritas. Denotamos esta correlación vectorial D con elementos d i sub>.
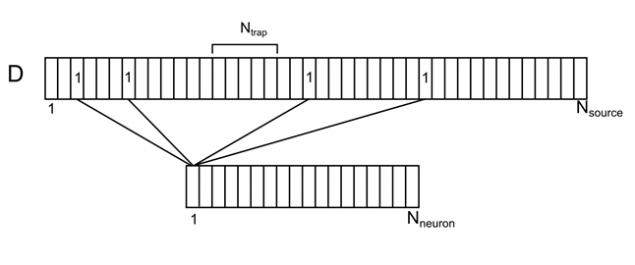
Figura 7. Correlación de las neuronas circundantes y sus contactos en
dendrita
Introducimos la notación:
N neurona sub> - número de neuronas que rodean
N fuente sub> - número de fuentes a una sola neurona
N trampa sub> - número de fuentes, creando un nivel de densidad de los neurotransmisores (trampa sináptica)
Ahora supongamos que algunas neuronas del entorno dan pico. Esto se puede tomar como una señal de que está disponible para ver nuestro neurona. Denotemos N sig sub> - el número de neuronas activas, la creación de una señal de información. Escribimos esta señal vector binario S.
Para todas las posiciones en la dendrita pero la más extrema, asumimos la densidad del mediador por la fórmula
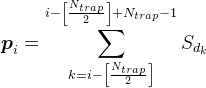
Por ejemplo, para que la señal se muestra en la figura siguiente, la densidad sináptica en la trampa de marcado es de 2 (la suma de las señales procedentes de la primera y cuarta de las neuronas).
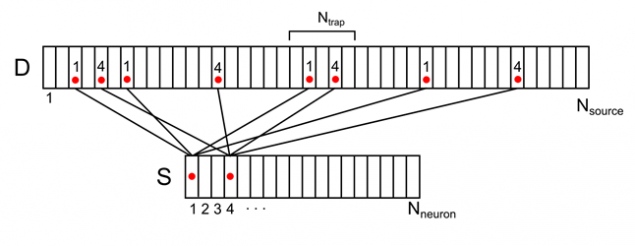
Figura 8. Visualización de la actividad de dos ambiente neuronal en el árbol dendrítico (sólo una parte de los bonos y la numeración)
Para cualquier señal arbitraria puede calcular la densidad del patrón de mediadores en la dendrita. Esta densidad oscilará entre 0 y N trampa sub>. El valor máximo se logrará si todas las fuentes estaban activos, formando trampa sináptica.
Utilizamos los valores medios de los parámetros característicos de la corteza de rata real (Braitenberg V., Schuz A., 1998). Sobre la base de estos, se obtienen los siguientes parámetros del modelo:
N neurona sub> = 650
N fuente sub> = 25000
N trampa sub> = 15
Suponemos que la señal se codifica por la actividad, por ejemplo, 1,5% de las neuronas de la corteza, mientras que
N sig sub> = 10
Es fácil de calcular la probabilidad de que una señal arbitraria que consiste en N sig sub> unidades, hay al menos un lugar en la dendrita, donde la densidad del mediador será exactamente K. Para un parámetro de probabilidad dada tener los valores siguientes (Tabla 2):
| K | P | ||||||||||||
| 0 | 0, 984 tr > | ||||||||||||
| 1 | 1 | ||||||||||||
| 2 | 1 | ||||||||||||
| 3 | 0, 996 | ||||||||||||
| 4 | 0, 287 | ||||||||||||
| 5 | 0, 016 | ||||||||||||
| 6 | 0, 001 | ||||||||||||
| 7 | 0 | ||||||||||||
| 8 | 0 | ||||||||||||
| 9 | 0 | ||||||||||||
| 10 | 0 | ||||||||||||
| ... Tabla 2. Tabla de la probabilidad de encontrar al menos una trampa con una densidad dada. La primera columna - el número necesario de fuentes activas en la trampa. En segundo lugar - la probabilidad de encontrar al menos un lugar de dendritas donde será un número de fuentes activas Es decir, para aquellos valores de los parámetros que están cerca de la configuración real de la corteza, para cualquier señal envolvente, afecta a alrededor del 1,5% de las neuronas, lo siguiente: • encontrar alrededor de 1,6% de las neuronas, que existe en las dendritas de la trampa, donde la señal cruza el 50% de los axones; • Casi todas las neuronas hay trampas, que intersecta al menos el 30% de los axones de señal. El significado de este resultado es muy interesante. Suponga que la información en la corteza es de alguna manera codificado actividad sincrónica de un número relativamente pequeño (N sig sub>) está dispuesto de forma compacta neuronas. No estamos hablando de toda la actividad del cerebro, y de los procesos de información en un pequeño volumen, donde las neuronas están numerados de 1 a nosotros N neurona sub>. Supongamos que el número de palabras de código S está acotado y forma una capacidad específica vocabulario T N dict sub>. Se puede calcular la probabilidad de que el mismo lugar "responder" de inmediato en dos señales. Los resultados de este cálculo para un diccionario de 10.000 señales en la tabla de abajo.
|
Tags
Vea también
Elon Musk cree que la inteligencia artificial es peligroso para la humanidad
La inteligencia artificial no es una amenaza para nosotros
El científico stuart russell acerca de la inteligencia artificial
10 películas sobre la inteligencia artificial, que debe mostrar cada
Arte en la inteligencia artificial
Inteligencia Artificial
¿Por qué estamos mal percibimos la inteligencia artificial
Steve wozniak no duda de la amenaza de la inteligencia artificial
















